
Reviews:
Microbial Cell, Vol. 7, No. 8, pp. 202 - 214; doi: 10.15698/mic2020.08.725
From the Uncharacterized Protein Family 0016 to the GDT1 family: Molecular insights into a newly-characterized family of cation secondary transporters

1 Louvain Institute of Biomolecular Science and Technology, UCLouvain, Louvain-la-Neuve, Belgium.
Keywords: UPF0016 family, Gdt1 family, Secondary transporters, Glycosylation, Calcium, Manganese, Gdt1p, TMEM165.
Received originally: 02/03/2020 Received in revised form: 25/05/2020
Accepted: 03/06/2020
Published: 15/06/2020
Correspondence:
Pierre Morsomme, Croix du Sud 4-5, Bte L7.07.14, B-1348 Louvain-la-Neuve, Belgium. Phone: (+32)10/47.26.23; pierre.morsomme@uclouvain.be
Conflict of interest statement: The authors declare no conflicts of interest.
Please cite this article as: Louise Thines, Jiri Stribny and Pierre Morsomme (2020). From the Uncharacterized Protein Family 0016 to the GDT1 family: Molecular insights into a newly-characterized family of cation secondary transporters. Microbial Cell 7(8): 202-214. doi: 10.15698/mic2020.08.725
Abstract
The Uncharacterized Protein Family 0016 (UPF0016) gathers poorly studied membrane proteins well conserved through evolution that possess one or two copies of the consensus motif Glu-x-Gly-Asp-(Arg/Lys)-(Ser/Thr). Members are found in many eukaryotes, bacteria and archaea. The interest for this protein family arose in 2012 when its human member TMEM165 was linked to the occurrence of Congenital Disorders of Glycosylation (CDGs) when harbouring specific mutations. Study of the UPF0016 family is undergone through the characterization of the bacterium Vibrio cholerae (MneA), cyanobacterium Synechocystis (SynPAM71), yeast Saccharomyces cerevisiae (Gdt1p), plant Arabidopsis thaliana (PAM71 and CMT1), and human (TMEM165) members. These proteins have all been identified as transporters of cations, more precisely of Mn2+, with an extra reported function in Ca2+ and/or H+ transport for some of them. Apart from glycosylation in humans, the UPF0016 members are required for lactation in humans, photosynthesis in plants and cyanobacteria, Ca2+ signaling in yeast, and Mn2+ homeostasis in the five aforementioned species. The requirement of the UPF0016 members for key physiological processes most likely derives from their transport activity at the Golgi membrane in human and yeast, the chloroplasts membranes in plants, the thylakoid and plasma membranes in cyanobacteria, and the cell membrane in bacteria. In the light of these studies on various UPF0016 members, this family is not considered as uncharacterized anymore and has been renamed the Gdt1 family according to the name of its S. cerevisiae member. This review aims at assembling and confronting the current knowledge in order to identify shared and distinct features in terms of transported molecules, mode of action, structure, etc., as well as to better understand their corresponding physiological roles.
INTRODUCTION
Proteins are classified into families in which members share a common evolutionary origin, reflecting their related functions and similarities in terms of sequence and/or structure. Therefore, when a novel protein is identified, its functional properties are often hypothesized based on the functional features of the group it is predicted to belong to. The number of defined protein families increased drastically with the development of genome sequencing. However, some of these families have absolutely no assigned function, for none of their members; these are the so-called Uncharacterized Protein Families (UPFs). This review focuses on one of these UPFs: the UPF0016 family (Pfam PF01169, TCDB number: 2.A.106). This protein family was described in 2014 as gathering membrane proteins found in many eukaryotes, bacteria, and archaea [1]. Numerous paralogs are mainly found in plants, which possess UPF0016 members in various subcellular compartments, including the Golgi [2], endoplasmic reticulum (ER) [2], and chloroplasts [3][4][5][6][7], whereas the non-plant eukaryotic UPF0016 members were all found at the Golgi up to now [8][9]. All UPF0016 members are defined by the presence of one or two copies of the Glu-φ-Gly-Asp-(Arg/Lys)-(Ser/Thr) consensus motif (with φ being any hydrophobic residue) [1]. Despite the high level of conservation within this protein family, some protein sub-groups share specific features in terms of length of the N-terminal part or enrichment in negatively charged residues in specific regions for instance [1]. From a broader view, the UPF0016 family belongs to the LysE superfamily that gathers eleven families of transport proteins that catalyze export of amino acids, lipids, and heavy metal ions [10].
–
Based on in silico approaches, the predicted topology of the eukaryotic UPF0016 members consists of two homologous clusters of three transmembrane spans with an opposite orientation in the membrane. These two clusters are separated by a central loop enriched in acidic amino acids (Figure 1) [1]. Interestingly, the predicted topology of the eukaryotic UPF0016 members is similar to that encountered in most of the families that belong to the LysE superfamily, mainly in terms of the presence of two internal repeats of three transmembrane spans [10]. More diversity in terms of topology can be observed among the prokaryotic members of the UPF0016 family. Indeed, some prokaryotes possess one gene coding for one UPF0016 member of three transmembrane spans that is predicted to auto-assemble as a homodimer while others possess two adjacent genes, encoding two different UPF0016 members of three transmembrane spans that are hypothesized to assemble as heterodimers. Finally, the eukaryotic-like topology, with six transmembrane spans, is also found among prokaryotes [1].
–
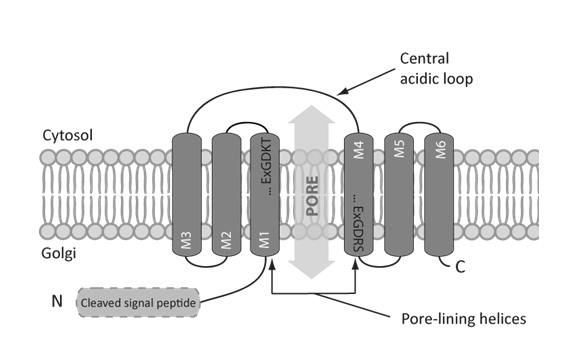 | FIGURE 1: Predicted topology of the eukaryotic UPF0016 members. The eukaryotic UPF0016 members are predicted to consist of two clusters of three transmembrane spans assembled in an antiparallel orientation and separated by a central loop enriched in negatively charged residues found at the cytosolic side in yeast, human, and bacteria. The featured UPF0016 motif Glu-φ-Gly-Asp-(Arg/Lys)-(Ser/Thr) is found in two copies, in the transmembrane spans one and four. Due to the hydrophilic nature of the residues found in this motif, and its localization in hydrophobic transmembrane spans, their two copies were hypothesized to form the pore of the protein through which the transported ions would cross the membrane [1]. |
–
When comparing the protein sequences of numerous UPF0016 members, the most conserved regions are the transmembrane spans and the featured motif Glu-φ-Gly-Asp-(Arg/Lys)-(Ser/Thr). This latter is predicted to localize in the transmembrane spans one and four of the eukaryotic members (Figure 1) [1]. Despite its hydrophobic environment, this motif contains two negatively charged (Asp and Glu), one positively charged (Arg or Lys), and one hydroxyl-containing (Ser or Thr) residue. Based on the role of similar residues in coordinating transported cations for proteins with available resolved structure [11], the two copies of the UPF0016 featured motif were hypothesized to form the pore of the putative transporter.
–
Since 2012 and the establishment of a causal link between the presence of mutations within TMEM165 (Transmembrane protein 165) and the occurrence of congenital disorders of glycosylation (CDGs) [8], many studies have been conducted on the functional properties of bacterial, yeast, plant, and human UPF0016 members. These studies reported a role of the UPF0016 members mainly in cation transport, thereby influencing essential processes like protein glycosylation in yeast and human, lactation in human, photosynthesis in plants and cyanobacteria, and Mn2+ toxicity in bacteria. This review aims at gathering the state-of-the-art knowledge accumulated on various UPF0016 members, thereby clarifying their molecular function in various organisms, and their subsequent implication in cellular processes. The mode of action (direction of transport, cations transported, structure-function relationship, etc.) of the UPF0016 members is also discussed in this review to shed light on conserved aspects but also on key differences that seem to occur within the family in terms of the aspects aforementioned, since this could be determinants of their physiological implication.
THE UPF0016 FAMILY AND CATION TRANSPORT
As previously stated, the UPF0016 family belongs to the LysE superfamily that gathers transporters of amino acids, lipids, and heavy metal ions [10]. In addition, similarities in terms of predicted topology of the UPF0016 members on the one hand, and of members of the cation-Ca2+ exchangers superfamily, that gathers transporters of Ca2+ against its electrochemical gradient by utilizing the downhill gradients of other cation species like H+, Na+, or K+[12], on the other hand, have also been observed. Based on these observations, the UPF0016 members were hypothesized to act as secondary cation transporters [1]. Since then, and as detailed below, several studies indicated transport of Mn2+ by all studied UPF0016 members, and of Ca2+ and/or protons by some of them.
–
Transport of Mn2+ by UPF0016 members
Since 2016, studies on the role of the six characterized UPF0016 members (MneA, SynPAM71, Gdt1p, PAM71, CMT1, and TMEM165) in Mn2+ homeostasis started to appear in the literature. In the plant Arabidopsis thaliana, the paralogs PAM71 (Photosynthesis-affected mutant 71, also called CCHA1, for Chloroplast Ca2+/H+ antiporter, or BICAT1, for Bivalent cation transporter 1) and CMT1 (Chloroplast Mn2+ transporter 1, also called BICAT2, for Bivalent cation transporter 2, or PAM71-HL) were both identified as involved in Mn2+ homeostasis from their localization at the thylakoid membrane [3][4][5] and inner envelope [4][6][7] of the chloroplasts, respectively. At the plant level, the two corresponding mutant plants show affected photosynthesis [3][4][5][6][7], reduced starch synthesis [4][5][6][7] and chlorophyll content [3][4][7], and decreased growth rate [3][4][5][6][7]. Interestingly, in both cases, the photosynthetic deficiency of the mutant plants could be partially suppressed in case of addition of Mn2+ to the growth medium [5][6][7]. Besides, these two chloroplast proteins are required for proper function and abundance of the Mn2+-containing oxygen-evolving complex of the photosystem II (PSII) (Mn4CaO5) [3][5][6][7] and both mutant plants show reduced incorporation of Mn2+ per unit of PSII [5][6]. Hence, these data all indicate a correlation between the photosynthetic deficiencies of the mutant plants and a reduced Mn2+ content within the chloroplasts. To further confirm their cation transport ability, CMT1 or PAM71 were individually expressed in the yeast strain pmr1Δ, devoid of its Golgi Ca2+-Mn2+ ATPase. This led in both cases to the suppression of the well-established Mn2+ sensitivity of pmr1Δ [5][6]. Expression of CMT1 in yeast further suppresses the growth defect observed under Mn2+-limited environment of the smf1Δ strain, devoid of its Mn2+ importer, through a re-increased cellular Mn2+ pool, again strengthening its ability to transport Mn2+[7]. Interestingly, the double mutant CMT1-PAM71 shows a phenotype closer to the CMT1 than that of the PAM71 mutant, thereby illustrating that CMT1 is the limiting step in Mn2+ delivery to the chloroplast. This means that CMT1 would function upstream of PAM71 for Mn2+ uptake over the chloroplast envelope and then the thylakoid membrane, respectively [6]. Taken together, these studies demonstrate the key role of both CMT1 and PAM71 in chloroplast development and Mn2+ supply to the PSII, which are both crucial for proper photosynthesis.
–
Similar to the observations in plants, the UPF0016 ortholog found in the cyanobacterium Synechocystis, SynPAM71 (also called Mnx, for Mn2+ exporter), is also crucial for maintenance of Mn2+ homeostasis and proper photosynthesis. The subcellular localization of SynPAM71 is not clearly established yet: Gandini et al. report localization both at the plasma membrane and, to a lesser extent, at the thylakoid membranes [13] whereas Brandenburg et al. localize SynPAM71 only at the thylakoid membranes [14]. The SynPAM71 loss-of-function line displays intracellular Mn2+ enrichment (particularly in the thylakoid lumen), reduced levels of chlorophyll, as well as reduced abundance of the PSII and defects in its photochemistry [13][14]. Interestingly, while Mn2+ supplementation to the external medium improves photosynthesis in the mutant plants, the cyanobacterial mutant shows a Mn2+-sensitive phenotype [13][14]. Additionally, the SynPAM71 mutant is not able to release previously-internalized radioactive 54Mn2+ from its internal pools, thereby suggesting toxic Mn2+ accumulation in this SynPAM71 loss-of-function line [14]. The transport ability of SynPAM71 was further supported by the fact that its expression in yeast also suppresses the Mn2+-sensitive phenotype of the pmr1Δ strain [14]. Taken together, these data all suggest a role of the Synechocystis UPF0016 member in Mn2+ export to prevent Mn2+ toxicity that could in turn impair essential processes like photosynthesis. Interestingly, similar effects on the photosynthetic efficiency were also attributed to the Chlamydomonas UPF0016 mutant GLD1 (Glucose-6-phosphate-1-dehydrogenase), thereby indicating a conserved function of the UPF0016 members from plants, algae, and cyanobacteria, in photosynthesis through regulation of the cation internal pools [5].
–
Recently, the MneA (Mn2+ exporter A) UPF0016 member from the bacterium Vibrio cholerae, a pathogenic gram-negative bacterium, was identified as a putative Mn2+ exporter. Indeed, the MneA mutant exhibits sensitivity to high Mn2+ concentration and shows an increased intracellular Mn2+ level when exposed to Mn2+, compared to a wild-type strain [15]. Due to the role of Mn2+ as antioxidant, the MneA mutant also shows increased resistance to H2O2 compared to the wild-type through internal accumulation of Mn2+[15]. Besides, expression of mneA suppresses the Mn2+-sensitive phenotype of an Escherichia coli strain carrying a mutation in its Mn2+ export gene mntP and decreases its Mn2+ content [15][16], thereby showing the function of MneA in Mn2+ export.
–
Last but not least, evidence regarding the involvement of the human UPF0016 member TMEM165 in Mn2+ homeostasis has also been made available. First, the abundance of a Golgi-localized protein whose stability is known to be Mn2+-sensitive, GPP130, is altered in TMEM165-depleted cells, thereby indicating a disturbed Mn2+ homeostasis at the Golgi level [17]. Besides, TMEM165 itself is rapidly and specifically degraded in lysosomes in response to excess of Mn2+ in the extracellular medium [18]. Glycosylation defects are known to occur in TMEM165-depleted cells (as will be further described below). Interestingly, addition of Mn2+ to the extracellular medium restores glycosylation in TMEM165-depleted cells [17]. In Saccharomyces cerevisiae, addition of Mn2+ to the yeast growth medium both triggers degradation of its Golgi-localized UPF0016 member Gdt1p (Gcr1-dependent translation factor) and suppresses the glycosylation defects that can be observed in gdt1Δ cells cultured in the presence of Ca2+ excess [17][19]. Besides, Gdt1p is involved in resistance to high Mn2+ concentrations, controls the cellular Mn2+ pools, and modulates the enzymatic activity of Sod2p, an enzyme that requires Mn2+ as cofactor for proper activity [20]. Interestingly, production of a truncated version of TMEM165 (lacking its first 78 N-terminal amino acids) in the yeast strain gdt1Δ restores its Mn2+ sensitivity and cellular Mn2+ stores, thereby illustrating a conserved function in Mn2+ homeostasis [21]. The transport activity of both TMEM165 (produced again with truncated 78 N-terminal amino acids) and Gdt1p was further demonstrated by producing these proteins in Fura-2-loaded Lactococcus lactis cells, in which a Mn2+-induced Gdt1p-dependent quenching of the fluorescence emitted by Fura-2 can be observed [20][21]. Taken together, these data clearly illustrate the key role of the UPF0016 members from various species in Mn2+ homeostasis.
–
Transport of Ca2+ by UPF0016 members
Apart from their role in Mn2+ homeostasis, the yeast, plant, and human UPF0016 members were suggested to be involved in Ca2+ homeostasis. Most of the evidence of the involvement of the UPF0016 members in Ca2+ homeostasis actually arises from studies conducted on the S. cerevisiae member Gdt1p. First of all, growth of the gdt1Δ strain is reduced in the presence of high external Ca2+ concentration (750 mM), compared to the wild-type, illustrating its role in coping with Ca2+ stress [9]. Interestingly, this increased sensitivity of the gdt1Δ strain towards Ca2+ could be suppressed when expressing (i) a truncated version of the human ortholog TMEM165 (lacking its first 78 N-terminal residues [21]), (ii) a truncated version of the A. thaliana UPF0016 members PAM71 (lacking its first 155 N-terminal residues [3]) or CMT1 (lacking its first 131 N-terminal residues [4]), or (iii) the Candida albicans ortholog CaGDT1[22], thereby suggesting a conserved function in Ca2+ homeostasis. Another indication that Gdt1p is involved in Ca2+ homeostasis lies in its established interaction with PMR1 at the genetic level. PMR1 encodes a well-characterized yeast Ca2+-Mn2+ P-type ATPase colocalizing with Gdt1p at the cis- and medial-Golgi. This genetic interaction is reflected by the fact that gdt1Δpmr1Δ is more sensitive to Ca2+ than both single mutants [19] and that GDT1 expression level and activity depends on PMR1 expression and ability to transport Ca2+ and/or Mn2+[23], thereby suggesting similar functions for these two proteins. Besides, the level of expression of GDT1 influences the cellular Ca2+ accumulation (most likely since Gdt1p modulates the intraluminal Golgi cation content and cations can exit the cell from the Golgi through secretory vesicles trafficking) as well as the Ca2+ response observed after exposure of yeast cells to a salt stress [19], all supporting its role in yeast Ca2+ homeostasis. In the yeast C. albicans, the comparison of the double deletion strain to each of the single deletions indicates that CaGdt1p and CaPmr1p also interact at the genetic level. Indeed, further deletion of CaGDT1 in a strain deleted for CaPMR1 increases (i) its sensitivity towards cell wall and ER stresses, (ii) its ability to accumulate radioactive 45Ca2+, and (iii) its hypersensitivity to inhibitors of the Ca2+-mediated calcineurin signaling pathway [24]. Transcriptomic analyses further revealed that CaGdt1p is involved in the regulation of cellular transport of metal ions and amino acids [24]. In this pathogen, CaGdt1p also interacts at the genetic level with the high-affinity plasma membrane Ca2+ channel CaCch1p/CaMid1p, again strengthening the role of CaGdt1p in Ca2+ homeostasis. Indeed, extra deletion of CaGDT1 in a strain deleted for CaCCH1 or CaMID1 suppresses its sensitivity to cold stress, but also increases its sensitivity to antifungal drugs [22].
–
Whole-cell patch-clamp analyses on HeLa cells overexpressing TMEM165 further enabled to assign a cation transport activity to TMEM165 [9]. Interestingly, the membrane currents that were observed for the TMEM165-expressing HeLa cells were decreased in the presence of the Ca2+-chelating agent EGTA, suggesting Ca2+ transport by TMEM165 [9]. The use of the Ca2+-sensitive fluorescent probe Fura-2 in HeLa cells overexpressing TMEM165 further supported the role of this human protein in Ca2+ homeostasis [9]. Like in yeast, TMEM165 genetically interacts with the human ortholog of Pmr1p, the Golgi Ca2+-Mn2+ ATPase SPCA1, in the way that the abundance of TMEM165 depends on the abundance and function of SPCA1. The authors of this study even suggest that SPCA1 and TMEM165 physically interact [25]. Interestingly, in mice, the level of expression of TMEM165 in lactating mammary tissues increases by a 25-fold factor during lactation while forced cessation of lactation leads to a rapid decrease of its expression. As about 40% of Ca2+ in milk (30-80 mM) is thought to be first stored in the Golgi lumen of the mammary epithelial cells to be then secreted into milk, both timing and magnitude of expression of TMEM165 place this protein as a potential contributor to mammary Golgi Ca2+ transport needs [26].
–
Finally, the two plant UPF0016 members from the A. thaliana chloroplasts, PAM71 and CMT1, were also linked to Ca2+ homeostasis, although their ability to transport Ca2+ cations still remains under debate. The identification of these two plant proteins as putative Ca2+ transporters arises from decreased radioactive 45Ca2+ uptake in isolated thylakoids and chloroplasts of the PAM71 and CMT1 mutant plants, respectively [4]. Besides, while modulation of the Ca2+ concentration in the chloroplast stroma is known to occur in the transition between light and dark phases of photosynthesis, this [Ca2+]stroma was detected as affected in the two mutant plants compared to the wild-type, thereby reflecting an influence on Ca2+ signaling [4]. In the light of the key role of Ca2+ in photosynthesis, both in terms of signaling and regulation of involved enzymes, the previously mentioned defects in terms of growth and photosynthesis observed in the two mutant plants could therefore partly be due to disturbed Ca2+ partitioning in the chloroplasts [3][4]. Further study of the thylakoid membrane PAM71 mutant plant shows that it is sensitive to high external Ca2+ concentrations and EGTA [3], and that it has a modified cytosolic Ca2+ content compared to the wild-type [3]. However, contradictory data regarding the role of the A. thaliana UPF0016 members in Ca2+ homeostasis are available: whereas Schneider et al. reported a higher Ca2+ uptake in the thylakoids of PAM71 mutants [5], Frank et al. observed a decrease in such Ca2+ uptake [4]. In addition, Frank et al. notified differences in terms of chloroplast Ca2+ content between wild-type and CMT1 mutant plants [4] whereas no such differences were observed by Zhang et al[7]. These contradictions reveal that further studies are needed to clearly evaluate the role of PAM71 and CMT1 in Ca2+ homeostasis, and to determine whether their impact on photosynthesis is of primary or secondary nature.
–
Recently, the Ca2+ transport activity of the yeast Gdt1p [19], the plant PAM71 [4], and the human TMEM165 [21] was further supported by the recording of a more pronounced Ca2+ influx in bacteria producing one of these UPF0016 members with the Ca2+-sensitive fluorescent probe Fura-2, compared to control cells containing an empty plasmid. Taken together, these data suggest that the Golgi UPF0016 members from human and yeast, and the chloroplast UPF0016 members from plants influence the cellular Ca2+ distribution through their transport ability.
–
At that stage, it is interesting to point out that, while Gdt1p, TMEM165, PAM71, and CMT1 have all been linked to both Ca2+ and Mn2+, there is no current indication of Ca2+ transport by the bacterial SynPAM71 and MneA. This might suggest that, within the UPF0016 protein family, some members could transport the two cations while others would transport only one or the other. Of course, this statement has to be confirmed by further biochemical characterization of these UPF0016 members.
–
Transport of protons by UPF0016 members
The UPF0016 members are hypothesized to work as secondary transporters. To date, few pieces of evidence indicate that the UPF0016 members would exchange Ca2+ and/or Mn2+ against H+ and would thereby be involved in pH homeostasis. In this context, Demaegd et al. first noticed a decreased lysosomal pH in both fibroblasts of TMEM165-deficient patients and TMEM165-depleted HeLa cells, in comparison with control individuals or cells [9]. More recently, the use of an in situ Golgi-localized pH-sensitive probe enabled to highlight acidification of the Golgi apparatus in the absence of TMEM165 [27]. In the same line, the A. thaliana PAM71 mutant plants show a pH-sensitive phenotype and an increased cytoplasmic pH, which is thought to reflect disturbed stromal pH whose proper maintenance is of importance for effective photosynthesis [3]. Nevertheless, a reduced H+-ATPase activity was also observed in PAM71 mutant plants [3][5]. Therefore, one cannot exclude that the effects of PAM71 on chloroplast pH homeostasis are of secondary nature rather than directly linked to H+ transport. In yeast, the Ca2+ transport activity of Gdt1p depends on the pH gradient across the Golgi membrane created by the Golgi proton V-ATPase. More precisely, Gdt1p seems to promote Ca2+ sequestration within the Golgi lumen when this organelle is correctly acidified by the V-ATPase, but works in the opposite direction, from the Golgi to the cytosol, when such acidification is disrupted by deleting the gene coding for the V-ATPase [28]. Interestingly, the Ca2+ influx observed in Fura-2-loaded L. lactis cells producing Gdt1p depends on the external pH in the way that the higher the external pH, the higher the influx rate, being consistent with the model of cation/proton antiporter [19]. Finally, since proton is known to be a by-product of all glycosylation reactions, including during milk production in the Golgi lumen of lactating mammary cells, the increased expression of TMEM165 during lactating periods may contribute to deacidify the Golgi lumen by transporting the accumulated protons to the cytosol in exchange of Mn2+ and/or Ca2+[26].
–
While these data all suggest a role of the human TMEM165, the yeast Gdt1p, and the A. thaliana PAM71 in pH homeostasis, there is still a lack of direct H+ transport evidence for the UPF0016 members. Therefore, to date, one cannot exclude that the altered pH homeostasis observed in case of disruption of the UPF0016 members could be a secondary consequence of altered Ca2+ and/or Mn2+ homeostasis.
–
Combining transport of Ca2+, Mn2+, and H+…
When combining the data aforementioned, it appears that bacterial, yeast, plant, and human UPF0016 members all transport Ca2+, Mn2+, and/or H+ as secondary transporters. Figure 2 illustrates the putative mechanisms of action of the studied UPF0016 members according to the biochemical data accumulated to now. Mainly, there is indirect evidence of transport of these three cations for the yeast Gdt1p and the human TMEM165. One of the possible mechanisms of transport with these three actors consists of a Ca2+-Mn2+/H+ antiporter, where Ca2+ and Mn2+ would be transported at the Golgi level in one direction in exchange of protons. However, and in accordance with the limited and relatively indirect evidence of transport of protons, it was suggested that these UPF0016 members would work as Ca2+/Mn2+ antiporters [23][29]. In A. thaliana, data from different studies led to the conclusion that Mn2+ cations and, if transported, Ca2+ cations would also be transported in the same direction, towards the chloroplast lumen [3][4][5][6][7], with indication of transport of protons only for PAM71 [3]. In cyanobacteria (SynPAM71) and bacteria (MneA), there is up to now only indication of Mn2+ transport. If working as secondary transporters, as suspected from their predicted topology, the nature of the counter-ion still remains to be identified. In other words, combination of the published data suggests putative differential specificity among the UPF0016 members. Further study of the substrate specificity of a wide range of UPF0016 members would answer this question and putatively lead to the establishment of a correlation between the substrate specificity of the UPF0016 members and evolution.
–
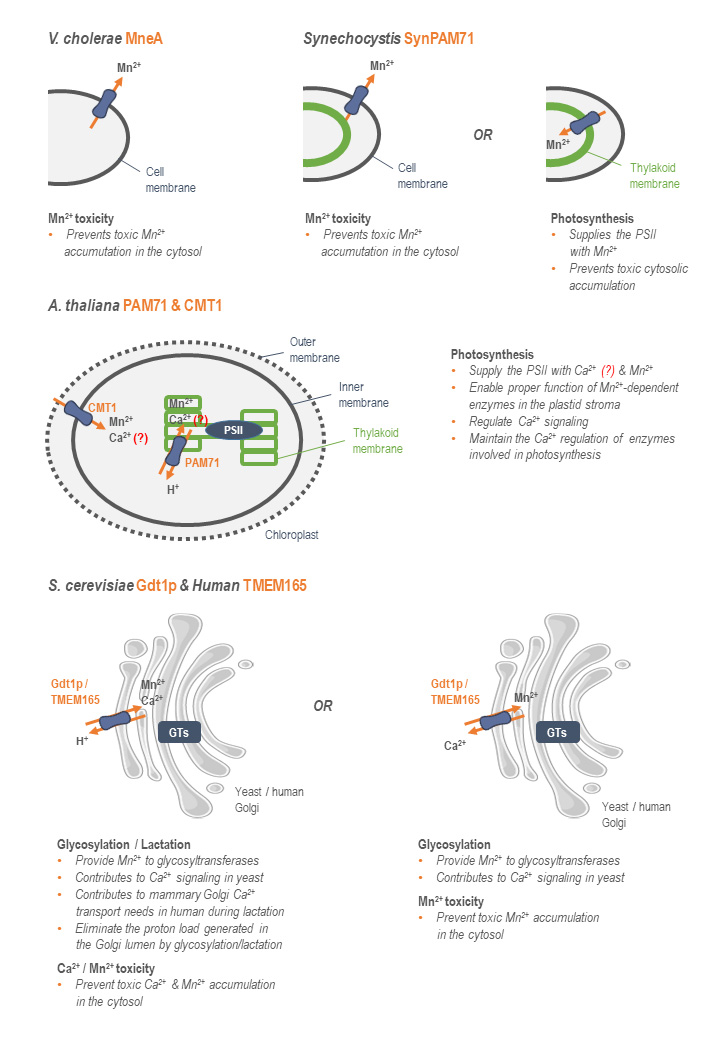 | FIGURE 2: Mode of action of the currently studied UPF0016 members. The direction of transport of the UPF0016 members' substrates from their respective subcellular localization is indicated by orange arrows. Their putative implication in key physiological processes is indicated. Functions that are still unclear in the literature are indicated by (?) (PSII: photosystem II, GTs: glycosyltransferases). |
–
Another missing piece of the puzzle is the direction of transport of these cations. Nevertheless, based on data interpretation, the human and yeast orthologs TMEM165 and Gdt1p could transport Ca2+ and Mn2+ from the cytosol to the Golgi lumen in exchange of protons from the Golgi to the cytosol, if working as Ca2+-Mn2+/H+ antiporters. In case of a Ca2+/Mn2+ antiporter, data suggest that Mn2+ would be imported in the Golgi in exchange of Ca2+[23][29]. In plants, Mn2+ and, if transported, Ca2+ would be transported from the cytosol to the stroma via CMT1, and then from the stroma to the thylakoid lumen via PAM71 [3][4][5][6][7]. In cyanobacteria, Mn2+ would be transported from the cytosol to the thylakoid lumen and/or the extracellular medium, depending on the considered subcellular localization of SynPAM71 [13][14]. Finally, the bacterial UPF0016 members are suspected as transporting Mn2+ from the cytosol to the extracellular medium [15][16], with no identified counter-ion. However, one should keep in mind that these directions of transport aforementioned are highly speculative and that the UPF0016 members, like numerous secondary transporters, could work reversely.
TRANSCRIPTIONAL AND TRANSLATIONAL REGULATION OF THE UPF0016 MEMBERS
In the light of their cation transport activity, it would not be surprising that the UPF0016 members would be regulated at the transcriptional and/or translational levels according to the concentrations of the transported substrates. In bacteria, 328 out of 333 screened strains that possess a UPF0016 member have the corresponding gene under the control of the Mn2+-regulated riboswitch yybP-ykoY, thereby indicating a transcriptional Mn2+-dependent regulation [16][30]. Additionally, five bacterial UPF0016 members (from the cyanobacterium Anabaena and four halobacteria) are under the control of the two-component system ManS (histidine kinase that senses the extracellular concentration of Mn2+)/ManR (DNA-binding response regulator that regulates expression of targeted genes) [31][32][33] that would modulate the expression of the UPF0016 members according to the neighboring Mn2+ concentration [34][35]. More direct expression analyses show that expression of the bacterial V. cholerae mneA is induced after addition of Mn2+ to the extracellular medium [15], while CMT1 expression in plant is reduced in the presence of Mn2+ excess [6]. Besides, the protein abundance of both Gdt1p and TMEM165 is reduced in case of external Mn2+ excess [18]. Although highly speculative, one might suggest based on these data that the presence of Mn2+ excess triggers (i) increased abundance of the bacterial UPF0016 members to detoxify the cytosol through their export activity, and (ii) decreased abundance of the eukaryotic UPF0016 members to prevent toxic Mn2+ excess in the lumen of the organelle this cation is sent to.
–
The activity of the yeast Gdt1p is reported to be negatively regulated by the Ca2+-induced calcineurin pathway [28]. No more is known regarding the Ca2+-dependent regulation of the UPF0016 members. This aspect should therefore be addressed in the near future.
THE UPF0016 FAMILY AND GLYCOSYLATION
Specific mutations in the gene coding for the human Golgi-localized UPF0016 member TMEM165 were identified as causing a sub-type of CDGs [8][36]. Since the scientific interest for the UPF0016 family mainly arises from this fact, this section gathers the up-to-date information that connects the family members to this essential cellular process. CDGs refer to a family of rare inherited diseases resulting in defects in the synthesis of glycans and in their attachment to proteins or lipids [37]. At that time, since the function of TMEM165 was undefined, the causal link between the presence of these mutations and the occurrence of the pathology was not fully understood. To date, five specific mutations within the gene coding for TMEM165 (four missense mutations: 126Arg>His, 126Arg>Cys, 304Gly>Arg, and 108Glu>Gly) and one mutation that activates a cryptic splice site (c.792+182 G>A)), identified in six patients, are reported to cause CDGs [8][36]. With more than 50% of eukaryotic proteins being glycosylated, it is not surprising that TMEM165-CDG patients show various and severe symptoms like growth and psychomotor retardation, muscular weakness, skeletal dysplasia, fat excess, and fever episodes among others, resulting in decreased life expectancy [8][36][38] (reviewed in [39]).
–
At the molecular level, patients suffering from TMEM165-CDGs, as well as TMEM165-depleted HeLa and HEK cells, show affected protein glycosylation profile, mainly in terms of relative hypo-sialylated and hypo-galactosylated N-glycans [8][17]. Interestingly, while these abnormalities point to defects in glycans synthesis at the Golgi, the Golgi network was found dilated and fragmented in affected individuals [8]. Besides, the glycosylation defects observed in culture cell lines could be suppressed by supplementing the growth medium with Mn2+[17], as previously stated, but also with galactose [40]. According to this observation, and based on the strong protein hypo-galactosylation in TMEM165-CDG patients, oral galactose supplementation was considered as a treatment of TMEM165-CDGs and has now proven to improve clinical and biochemical parameters of the patients [40]. Antisense RNA therapy targeting TMEM165 mRNAs was also successfully considered in case of pathogenic splicing (mutation c.792+182 G>A) [41].
–
The role of Gdt1p, the S. cerevisiae Golgi-localized UPF0016 member, in glycosylation efficiency has also been investigated. Surprisingly, no glycosylation deficiency could be detected in the strain deleted for GDT1. Protein glycosylation is however affected when growing the gdt1Δ strain in a Ca2+-rich medium (500 mM) [17][19]. Similar to observations in human cells, glycosylation is restored upon the additional presence of Mn2+ cations [17][19]. Structural analysis of the yeast glycans, mainly constituted of polymannan chains, further revealed that the gdt1Δ mutant cultured in presence of high Ca2+ concentrations presents strong late Golgi glycosylation defects with a lack of α-1,2 mannoses substitution and α-1,3 mannoses termination [23].
–
Finally, it was shown in zebrafish that inhibition of TMEM165 expression in developing embryos causes altered initiation, processing, and extension of N-glycans, together with altered cartilage and bone development [42]. Taken together, these data indicate that, despite not being identified as direct actors in glycosylation, the UPF0016 members are crucial for proper protein glycosylation.
FROM ALTERED UPF0016 TRANSPORT FUNCTION TO DISTURBED PHYSIOLOGICAL PROCESSES
The previous sections clearly illustrate that the UPF0016 members influence the cellular cation distribution, most likely through their common ability to transport Mn2+ but also Ca2+ and/or H+ for some of them. Disruption of the ionic intracellular pools in case of malfunction of the UPF0016 members might in turn influence essential physiological processes in their respective organism, as further detailed in this section. Of course, while focused on the UPF0016 members, it is important to mention that these transporters most likely act in concert with other transporters from other protein families that are also essential for proper cation homeostasis.
–
The link between TMEM165 and glycosylation can first be examined in the light of its transport activity. Many glycosyltransferases involved in glycosylation actually require interaction with Mn2+ as cofactor for proper activity. This has for instance been directly demonstrated for the enzymes α-1,3-N-acetylgalactosaminyltransferase, α-1,3-galactosyltransferase, β-1,3-glucuronosyltransferase, and β-1,4-galactosyltransferase among others [43][44][45]. Interestingly, the glycosylation defects observed in TMEM165-CDG patients mainly correspond to altered activity of the Golgi β-1,4-galactosyltransferase that requires Mn2+ cations as cofactor for proper activity [17]. Therefore, the glycosylation defects observed in TMEM165-CDG patients might arise from Golgi Mn2+ disturbances, which would in turn impair the enzymatic activity of this specific glycosyltransferase. Similarly, some S. cerevisiae glycosyltransferases also require Mn2+ as cofactor: Och1p, Mnn1p, Mnn2p, Mnn5p, and Mnn9p [46][47][48][49]. The glycosylation defects observed in the yeast gdt1Δ strain grown in the presence of Ca2+ actually correspond to altered activity of the Mn2+-dependent mannosyltransferases Mnn1p, Mnn2p, and Mnn5p [23], suggesting again that these glycosylation defects might derive from altered Mn2+ content within the Golgi. Nevertheless, as TMEM165 and Gdt1p both seem to transport also Ca2+ at the Golgi membrane, one cannot rule out the possibility that the glycosylation defects are also partly caused by a disturbed Ca2+ balance within the Golgi lumen (and/or cytosol). Indeed, Ca2+ cations are essential for proper vesicular trafficking through their role in membrane fusion, and for activity and stability of enzymes involved in glycosylation [50][51][52]. Apart from an implication in glycosylation, transport of Ca2+ by Gdt1p most likely modulates cellular Ca2+-induced signaling in yeast (since the modulation of the cytosolic Ca2+ concentration observed after a salt stress is affected in case of deletion of GDT1) [19]. Finally, transport of Ca2+ and/or Mn2+ cations in the Golgi lumen by the UPF0016 members might constitute a detoxification pathway to prevent toxic accumulation in the cytosol in case of cation excess. Indeed, once inside the Golgi lumen, these ions can be directed out of the cell through the secretory pathway.
–
Apart from providing the secretory pathway with Ca2+ and Mn2+, TMEM165 and Gdt1p are also suggested to modulate the pH homeostasis of the Golgi. According to this latter statement, Gdt1p would be a putative actor in eliminating the proton load generated by glycosylation, in exchange of transport of divalent cations from the cytosol to the Golgi [28]. In the same line, the increased expression of TMEM165 during lactation might reflect its role in providing the Golgi lumen of lactating mammary cells with Ca2+ as nutrient and Mn2+ as cofactor of enzymes, but also in removing the protons generated as by-product of lactose production out of the Golgi lumen [26][53]. Since there are no clear proton export mechanisms from the Golgi to now both in yeast and human, this involvement of the UPF0016 members in Golgi pH homeostasis appears as highly interesting for further investigation in the near future.
–
Before the characterization of the plant chloroplast UPF0016 members CMT1 and PAM71, the mechanism by which Mn2+ and Ca2+ cations were delivered into the chloroplast lumen remained elusive. The current model for Mn2+, and putatively for Ca2+ delivery in the chloroplast of A. thaliana is that these cations would first be transported from the cytosol into the chloroplast stroma through the inner envelope CMT1, and further transferred to the thylakoid lumen by PAM71 [7]. The photosynthetic defects observed in case of malfunction of these proteins most likely arise from disturbed Mn2+ content within the chloroplasts. Indeed, Mn2+ is a structural component of the PSII. In photosynthetic organisms, the PSII mediates splitting of water into oxygen, protons, and electrons. While O2 is released, the PSII thereby provides electrons for further photosynthetic reactions and generates a proton gradient used by the ATP synthase to generate ATP. Affected function of the PSII therefore leads to less reducing power and ATP synthesis [54], which in turn affects plant growth and development, as observed in the UPF0016 mutant plants (reviewed in [55]). In addition, both PAM71 and CMT1 most likely play a role in maintaining proper activity of Mn2+-requiring enzymes in the plastid stroma. As illustration, Mn2+ cations are known to activate the rubisco in the chloroplasts [56]. Besides, CMT1 and PAM71 influence Ca2+ homeostasis from their localization at the chloroplast, which could in turn also impair photosynthesis. Indeed, Ca2+ is a regulator of chloroplast enzymes, and proper modulation of the Ca2+ concentration within the chloroplast is essential for signaling during photosynthesis [4]. On the other hand, A. thaliana additionally possesses two ER-localized (PML4 and PML5) and one Golgi-localized (PML3) UPF0016 members [2]. The putative role in glycosylation of these UPF0016 members at the plant secretory pathway still remains to be investigated, so are their substrate specificity and their overall role in plant cell physiology.
–
Since the subcellular localization of the cyanobacterial SynPAM71 is still under debate (both at the plasma membrane and thylakoid membranes, suggesting dual targeting, according to [13], or only at the thylakoid membrane according to [14]), its physiological role has to be examined in the light of these two possibilities. At the thylakoid membrane, SynPAM71 would have similar functions than in plants, i.e. contributing to Mn2+ supply to the Mn4CaO5 oxygen-evolution center, but also in sequestering Mn2+ within the thylakoid lumen to prevent toxic accumulation in the cytosol. At the plasma membrane, this protein would balance the otherwise harmful effects of Mn2+ accumulation in the cytoplasm, hence preventing Mn2+ toxicity. In this context, the photosynthetic deficiency observed in the mutant line would derive from reduced Mn2+ bioavailability for the PSII, and/or from toxic cytosolic Mn2+ accumulation [13][14]. Determination of the exact subcellular localization of SynPAM71 thereby appears as essential to determine its physiological role.
–
Finally, in bacteria, the UPF0016 members would help maintaining physiological Mn2+ concentrations within the cytosol from their localization at the cell membrane, most likely with a Mn2+-dependent regulation to cope with a wide range of environmental stresses. Besides, Mn2+ is known to be essential for virulence of pathogenic bacteria [57]. Indeed, due to the requirement of Mn2+-dependent enzymes for virulence, bacteria must possess functional and well-regulated Mn2+ transporters. Maintenance of bacterial Mn2+ homeostasis is even more important in the light of the fact that mammalian host cells have developed strategies to sequester Mn2+ from bacterial invaders as a defense mechanism [57][58]. Therefore, it would be highly interesting to investigate the importance of the bacterial UPF0016 members during interactions with eukaryotic hosts for example.
STRUCTURE-FUNCITON RELATIONSHIP OF THE UPF0016 MEMBERS
Alignment of the protein sequences of the UPF0016 members reveals that the most conserved regions are the two copies of the featured motif Glu-φ-Gly-Asp-(Arg/Lys)-(Ser/Thr) (Figure 3). Therefore, the structure-function relationship of the UPF0016 members was analyzed mainly in terms of importance of the residues that constitute the two motifs to evaluate their putative role in ion coordination, but also of the residues that are found mutated in the TMEM165-CDG patients to gain insight into the mechanism of pathogenicity. Table 1 provides an overview of the mutations that have an effect on protein function, stability, or subcellular localization.
–
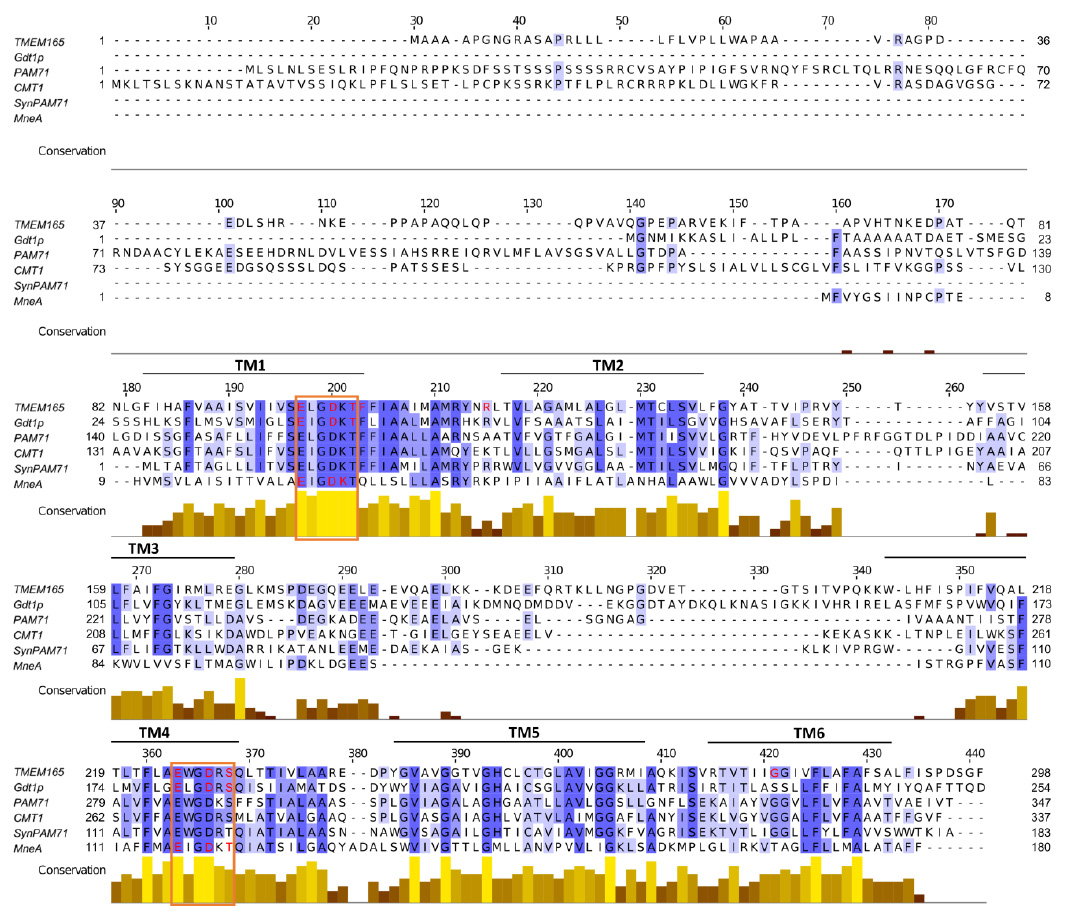 | FIGURE 3: Multiple alignment of the amino acid sequences of the studied UPF0016 members. The alignment was carried out using Clustal Omega and visualized in Jalview. Residues are shaded in blue depending on their conservation. The conservation histogram provides quantitative evaluation of the degree of conservation of the physico-chemical properties of the amino acids at the corresponding position. Putative transmembrane spans predicted by TMAP are indicated above the sequences (TM1 – TM6). The highly conserved UPF0016 motifs are framed in orange. Residues identified as altering the protein stability, localization, and/or activity are highlighted in red. |
–
First, due to the presence of negatively charged (Asp and Glu), positively charged (Lys or Arg), and hydroxyl-containing residues (Thr or Ser) within the UPF0016 motif predicted to be incorporated in hydrophobic transmembrane spans, these two copies were perceived as good candidates for the formation of the pore for cation transport. In yeast, production of Gdt1p with mutations in Ala of the acidic (53Glu & 204Glu, and 56Asp & 207Asp) and polar uncharged (58Thr and 209Ser) residues of the motifs in the gdt1Δpmr1Δ strain, that is sensitive to high Ca2+ concentrations and shows impaired Ca2+ response to a salt stress, failed to suppress these defects, whereas non-mutated Gdt1p and the other mutated versions from the motif did [59]. In the same line, expression of the mutated versions of GDT1 coding for proteins with mutation of the acidic residues of the conserved motifs (53Glu & 204Glu, and 56Asp & 207Asp) in the gdt1Δ strain fails to suppress the glycosylation defects observed in the presence of high Ca2+ concentration [23]. Altogether, this suggests that these residues are essential for the Ca2+ transport activity of Gdt1p (given that these mutated proteins are produced and well localized as verified by [59]). In human, the ability of wild-type or mutated TMEM165 to rescue glycosylation defects observed in TMEM165-KO cells was similarly used to investigate the involvement of the residues of the conserved motifs, as well as of flanking residues, in protein functionality [29]. Interestingly, the mutated proteins from the motifs 248Glu, 111Asp & 251Asp, 113Thr, and 253Ser, as well as from the flanking residue 114Phe, were all unable to restore glycosylation. While none of these mutations affected protein stability, the mutated proteins 251Asp and 253Ser were found mislocalized in vesicular structures throughout the cytoplasm. Finally, the TMEM165 mutated proteins from the motifs 108Glu, 111Asp & 251Asp, 113Thr, and 253Ser are resistant to Mn2+-induced degradation, in contrast to the wild-type TMEM165 that is known to be degraded in the presence of high Mn2+ concentrations [29]. These data clearly illustrate the role of the acidic and polar uncharged residues of the conserved motifs in activity, localization, and Mn2+-induced sensitivity of TMEM165. Finally, the mutated forms 19Glu & 116Glu, 22Asp & 119Asp, 23Lys, and 121Thr in Ala from the conserved motifs of the bacterial MneA are unable to complement the Mn2+ sensitivity of an E. coli mutant strain devoid of its Mn2+ exported MntP [16], thereby illustrating their key importance for proper protein function in Mn2+ homeostasis. Taken together, study of the primary structure-function relationship of the UPF0016 members from yeast, human, and bacteria illustrates a conserved importance of the residues that constitute the UPF0016 motifs, especially of the acidic and polar uncharged ones, for protein function, as well as an additional role in protein localization and Mn2+ sensitivity in human. More advanced structural analyses are, however, required to confirm the implication of these residues in cation coordination at the predicted pore of the transporter.
–
TABLE 1. Summary of the mutations within the UPF0016 members MneA (V. cholerae), Gdt1p (S. cerevisiae), and TMEM165 (human) that have an effect on their transport function, stability, or subcellular localization. |
 |
| [16][21][23][29][59][60] |
–
The effect of the naturally-occurring mutations among TMEM165-CDG patients were also examined in terms of protein localization, expression, and activity. In this context, the mutation c.792+182 G>A (that activates a cryptic splice site) was the only one identified as conferring protein instability [60]. In contrast, the mutations 126Arg>His, 126Arg>Cys, and 304Gly>Arg affect the Golgi subcellular localization of TMEM165. Interestingly, the mutations corresponding to 126Arg>His and 126Arg>Cys were also introduced in GDT1 and expressed in the gdt1Δ yeast strain (no residue equivalent to 304Gly being found in the yeast Gdt1p). Since their expression restores the ability of gdt1Δ to grow in Ca2+-rich medium, these proteins seem to be functional, thereby suggesting that the pathogenicity of these mutations is linked to altered localization rather than deficient protein activity [60]. More recently, due to optimization of the production of TMEM165 in the yeast gdt1Δ strain, its mutated versions were produced in this host. Using this yeast model of the disease, combined with transport assays carried out in TMEM165-producing L. lactis cells loaded with the fluorescent probe Fura-2, it appears that the mutation 108Glu>Gly leads to decreased protein activity while such reduced activity is not clear for the other mutants [21]. Altogether, these results illustrate that the pathogenicity of the mutations arises from distinct mechanisms than can now be unraveled through the biochemical tools aforementioned: altered expression (c.792+182 G>A), altered localization (126Arg>His, 126Arg>Cys, and 304Gly>Arg), or altered transport activity (108Glu>Gly) [21][60].
CONCLUSION
Molecular characterization of the UPF0016 members tremendously increased since their identification in 2012. The implication of the UPF0016 members in regulating cation homeostasis, thereby influencing various essential processes, is now well established in diverse organisms. For this reason, the UPF0016 family has been recently renamed as the Gdt1 family, according to the name of the yeast ortholog whose study significantly contributed to the characterization of this protein family (UniProt). Nevertheless, several aspects related to the UPF0016 family still remain to be investigated. Development of direct transport assays for the UPF0016 members in their endogenous host or proteoliposomes appears as one of the key approaches to confirm the identity of the transported substrates and their direction of transport. High-resolution structural analysis of the UPF0016 members also appears as essential for a better understanding of their mode of action.
–
Besides, recent results suggest that the spectrum of action of the Gdt1 family could be broader than what is currently known. Indeed, TMEM165 was found overexpressed in hepatocellular carcinoma and involved in cancer invasive activity, through a mechanism that still needs to be uncovered [61]. Another example lies in the recent identification of TMEM165 splice variants that localize at the ER, rather than at the Golgi, which also raises the possibility of additional function of TMEM165 that have not been identified yet [62]. Further studies of the Gdt1 family members will therefore most likely unravel new aspects on this captivating family of secondary transporters.
REFERENCES
- Demaegd D, Colinet AS, Deschamps A, Morsomme P (2014). Molecular evolution of a novel family of putative calcium transporters. PLoS ONE 9(6). 10.1371/journal.pone.0100851
- Hoecker N, Honke A, Frey K, Leister D, Schneider A (2020). Homologous Proteins of the Manganese Transporter PAM71 Are Localized in the Golgi Apparatus and Endoplasmic Reticulum. Plants 9(2): 239. 10.3390/plants9020239
- Wang C, Xu W, Jin H, Zhang T, Lai J, Zhou X, Zhang S, Liu S, Duan X, Wang H, Peng C, Yang C (2016). A Putative Chloroplast-Localized Ca2+/H+ Antiporter CCHA1 Is Involved in Calcium and pH Homeostasis and Required for PSII Function in Arabidopsis. Mol Plant 9(8): 1183-1196. 10.1016/j.molp.2016.05.015
- Frank J, Happeck R, Meier B, Hoang MTT, Stribny J, Hause G, Ding H, Morsomme P, Baginsky S, Peiter E (2019). Chloroplast-localized BICAT proteins shape stromal calcium signals and are required for efficient photosynthesis. New Phytol 221(2): 866-880. 10.1111/nph.15407
- Schneider A, Steinberger I, Herdean A, Gandini C, Eisenhut M, Kurz S, Morper A, Hoecker N, Rühle T, Labs M, Flügge UI, Geimer S, Schmidt SB, Husted S, Weber APM, Spetea C, Leister D (2016). The evolutionarily conserved protein PHOTOSYNTHESIS AFFECTED MUTANT71 is required for efficient manganese uptake at the thylakoid membrane in Arabidopsis. Plant Cell 28(4): 892-910. 10.1105/tpc.15.00812
- Eisenhut M, Hoecker N, Schmidt SB, Basgaran RM, Flachbart S, Jahns P, Eser T, Geimer S, Husted S, Weber APM, Leister D, Schneider A (2018). The Plastid Envelope CHLOROPLAST MANGANESE TRANSPORTER1 Is Essential for Manganese Homeostasis in Arabidopsis. Mol Plant 11(7): 955-969. 10.1016/j.molp.2018.04.008
- Zhang B, Zhang C, Liu C, Jing Y, Wang Y, Jin L, Yang L, Fu A, Shi J, Zhao F, Lan W, Luan S (2018). Inner Envelope CHLOROPLAST MANGANESE TRANSPORTER 1 Supports Manganese Homeostasis and Phototrophic Growth in Arabidopsis. Mol Plant 11(7): 943-954. 10.1016/j.molp.2018.04.007
- Foulquier F, Amyere M, Jaeken J, Zeevaert R, Schollen E, Race V, Bammens R, Morelle W, Rosnoblet C, Legrand D, Demaegd D, Buist N, Cheillan D, Guffon N, Morsomme P, Annaert W, Freeze HH, Van Schaftingen E, Vikkula M, Matthijs G (2012). TMEM165 deficiency causes a congenital disorder of glycosylation. Am J Hum Genet 91(1): 15-26. 10.1016/j.ajhg.2012.05.002
- Demaegd D, Foulquier F, Colinet AS, Gremillon L, Legrand D, Mariot P, Peiter E, Van Schaftingen E, Matthijs G, Morsomme P (2013). Newly characterized Golgi-localized family of proteins is involved in calcium and pH homeostasis in yeast and human cells. Proc Natl Acad Sci U S A 110(17): 6859-6864. 10.1073/pnas.1219871110
- Tsu BV, Saier MH, Jr. (2015). The LysE Superfamily of Transport Proteins Involved in Cell Physiology and Pathogenesis. PLOS ONE 10(10): e0137184. 10.1371/journal.pone.0137184
- Waight AB, Pedersen BP, Schlessinger A, Bonomi M, Chau BH, Roe-Zurz Z, Risenmay AJ, Sali A, Stroud RM (2013). Structural basis for alternating access of a eukaryotic calcium/proton exchanger. Nature 499(7456): 107-110. 10.1038/nature12233
- Cai X, Lytton J (2004). The Cation/Ca2+ Exchanger Superfamily: Phylogenetic Analysis and Structural Implications. Mol Biol Evol 21(9): 1692–1703. 10.1093/molbev/msh177
- Gandini C, Schmidt SB, Husted S, Schneider A, Leister D (2017). The transporter SynPAM71 is located in the plasma membrane and thylakoids, and mediates manganese tolerance in Synechocystis PCC6803. New Phytol 215(1): 256-268. 10.1111/nph.14526
- Brandenburg F, Schoffman H, Kurz S, Krämer U, Keren N, Weber APM, Eisenhut M (2017). The Synechocystis manganese exporter mnx is essential for manganese homeostasis in cyanobacteria. Plant Physiol 173(3): 1798-1810. 10.1104/pp.16.01895
- Fisher CR, Wyckoff EE, Peng ED, Payne SM (2016). Identification and Characterization of a Putative Manganese Export Protein in Vibrio cholerae. J Bacteriol 198(20): 2810-2817. 10.1128/jb.00215-16
- Zeinert R, Martinez E, Schmitz J, Senn K, Usman B, Anantharaman V, Aravind L, Waters LS (2018). Structure-function analysis of manganese exporter proteins across bacteria. J Biol Chem 293(15): 5715-5730. 10.1074/jbc.M117.790717
- Potelle S, Morelle W, Dulary E, Duvet S, Vicogne D, Spriet C, Krzewinski-Recchi MA, Morsomme P, Jaeken J, Matthijs G, De Bettignies G, Foulquier F (2016). Glycosylation abnormalities in Gdt1p/TMEM165 deficient cells result from a defect in Golgi manganese homeostasis. Hum Mol Genet 25(8): 1489-1500. 10.1093/hmg/ddw026
- Potelle S, Dulary E, Climer L, Duvet S, Morelle W, Vicogne D, Lebredonchel E, Houdou M, Spriet C, Krzewinski-Recchi MA, Peanne R, Klein A, De Bettignies G, Morsomme P, Matthijs G, Marquardt T, Lupashin V, Foulquier F (2017). Manganese-induced turnover of TMEM165. Biochem J 474(9): 1481-1493. 10.1042/BCJ20160910
- Colinet AS, Sengottaiyan P, Deschamps A, Colsoul ML, Thines L, Demaegd D, Duchêne MC, Foulquier F, Hols P, Morsomme P (2016). Yeast Gdt1 is a Golgi-localized calcium transporter required for stress-induced calcium signaling and protein glycosylation. Sci Rep 6: 24282. 10.1038/srep24282
- Thines L, Deschamps A, Sengottaiyan P, Savel O, Stribny J, Morsomme P (2018). The yeast protein Gdt1p transports Mn2+ ions and thereby regulates manganese homeostasis in the Golgi. J Biol Chem 293(21): 8048-8055. 10.1074/jbc.RA118.002324
- Stribny J, Thines L, Deschamps A, Goffin P, Morsomme P (2020). The human Golgi protein TMEM165 transports calcium and manganese in yeast and bacterial cells. J Biol Chem 295(12):3865-3874. 10.1074/jbc.RA119.012249
- Wang Y, Wang J, Cheng J, Xu D, Jiang L (2015). Genetic interactions between the Golgi Ca2+/H+ exchanger Gdt1 and the plasma membrane calcium channel Cch1/Mid1 in the regulation of calcium homeostasis, stress response and virulence in Candida albicans. FEMS Yeast Res 15(7): fov069-fov069. 10.1093/femsyr/fov069
- Dulary E, Yu S-Y, Houdou M, de Bettignies G, Decool V, Potelle S, Duvet S, Krzewinski-Recchi M-A, Garat A, Matthijs G, Guerardel Y, Foulquier F (2017). Investigating the function of Gdt1p in yeast Golgi glycosylation. Biochim Biophys Acta 1862(3):394-402. 10.1016/j.bbagen.2017.11.006
- Jiang L, Wang J, Asghar F, Snyder N, Cunningham KW (2018). CaGdt1 plays a compensatory role for the calcium pump CaPmr1 in the regulation of calcium signaling and cell wall integrity signaling in Candida albicans. Cell Commun Signal 16(1): 33. 10.1186/s12964-018-0246-x
- Lebredonchel E, Houdou M, Hoffmann HH, Kondratska K, Krzewinski MA, Vicogne D, Rice CM, Klein A, Foulquier F (2019). Investigating the functional link between TMEM165 and SPCA1. Biochem J 476(21): 3281-3293. 10.1042/bcj20190488
- Reinhardt TA, Lippolis JD, Sacco RE (2014). The Ca2+/H+ antiporter TMEM165 expression, localization in the developing, lactating and involuting mammary gland parallels the secretory pathway Ca2+ ATPase SPCA1. Biochem Biophys Res Commun 445(2): 417-421. 10.1016/j.bbrc.2014.02.020
- Wang H, Yang Y, Huang F, He Z, Li P, Zhang W, Zhang W, Tang B (2020). In situ Fluorescent and Photoacoustic Imaging of Golgi pH to Elucidate the Function of Transmembrane Protein 165. Anal Chem 92(4):3103-3110. 10.1021/acs.analchem.9b04709
- Snyder NA, Stefan CP, Soroudi CT, Kim A, Evangelista C, Cunningham KW (2017). H+ and Pi Byproducts of Glycosylation Affect Ca2+ Homeostasis and Are Retrieved from the Golgi Complex by Homologs of TMEM165 and XPR1. G3 7(12): 3913-3924. 10.1534/g3.117.300339
- Lebredonchel E, Houdou M, Potelle S, de Bettignies G, Schulz C, Krzewinski Recchi M-A, Lupashin V, Legrand D, Klein A, Foulquier F (2019). Dissection of TMEM165 function in Golgi glycosylation and its Mn2+ sensitivity. Biochimie 165: 123-130. 10.1016/j.biochi.2019.07.016
- Dambach M, Sandoval M, Updegrove Taylor B, Anantharaman V, Aravind L, Waters Lauren S, Storz G (2015). The Ubiquitous yybP-ykoY Riboswitch Is a Manganese-Responsive Regulatory Element. Mol Cell 57(6): 1099-1109. 10.1016/j.molcel.2015.01.035
- Ogawa T, Bao DH, Katoh H, Shibata M, Pakrasi HB, Bhattacharyya-Pakrasi M (2002). A two-component signal transduction pathway regulates manganese homeostasis in Synechocystis 6803, a photosynthetic organism. J. Biol. Chem277(32): 28981–28986. 10.1074/jbc.M204175200
- Yamaguchi K, Suzuki I, Yamamoto H, Lyukevich A, Bodrova I, Los D-A, Piven I, Zinchenko V, Kanehisa M, Murata N (2002). A two-component Mn2+-sensing system negatively regulates expression of the mntCAB operon in Synechocystis. Plant Cell 14(11): 2901–2913. 10.1105/tpc.006262
- Zorina A, Sinetova MA, Kupriyanova EV, Mironov KS, Molkova I, Nazarenko LV, Zinchenko V, Los DA (2016). Synechocystis mutants defective in manganese uptake regulatory system, ManSR, are hypersensitive to strong light. Photosynth Res 130(1-3): 11–17. 10.1007/s11120-015-0214-x
- Huang W, Wu Q-Y (2004). Identification of genes controlled by the manganese response regulator, ManR, in the cyanobacterium, Anabaena sp. PCC 7120. Biotechnol Lett 26(18): 1397-1401. 10.1023/B:BILE.0000045639.26579.6f
- Leyn SA, Rodionov DA (2015). Comparative Genomics of DtxR Family Regulons for Metal Homeostasis in Archaea. J Bacteriol 197(3): 451. 10.1128/JB.02386-14
- Schulte Althoff S, Gruneberg M, Reunert J, Park JH, Rust S, Muhlhausen C, Wada Y, Santer R, Marquardt T (2016). TMEM165 Deficiency: Postnatal Changes in Glycosylation. JIMD Rep 26: 21-29. 10.1007/8904_2015_455
- Jaeken J, Matthijs G (2007). Congenital disorders of glycosylation: a rapidly expanding disease family. Annu Rev Genomics Hum Genet 8: 261-278. 10.1146/annurev.genom.8.080706.092327
- Zeevaert R, de Zegher F, Sturiale L, Garozzo D, Smet M, Moens M, Matthijs G, Jaeken J (2013). Bone Dysplasia as a Key Feature in Three Patients with a Novel Congenital Disorder of Glycosylation (CDG) Type II Due to a Deep Intronic Splice Mutation in TMEM165. JIMD Rep 8: 145-152. 10.1007/8904_2012_172
- Dulary E, Potelle S, Legrand D, Foulquier F (2017). TMEM165 deficiencies in Congenital Disorders of Glycosylation type II (CDG-II): Clues and evidences for roles of the protein in Golgi functions and ion homeostasis. Tissue Cell 49: 150-156. 10.1016/j.tice.2016.06.006
- Morelle W, Potelle S, Witters P, Wong S, Climer L, Lupashin V, Matthijs G, Gadomski T, Jaeken J, Cassiman D, Morava E, Foulquier F (2017). Galactose Supplementation in Patients With TMEM165-CDG Rescues the Glycosylation Defects. J Clin Endocrinol Metab 102(4): 1375-1386. 10.1210/jc.2016-3443
- Yuste-Checa P, Medrano C, Gamez A, Desviat LR, Matthijs G, Ugarte M, Perez-Cerda C, Perez B (2015). Antisense-mediated therapeutic pseudoexon skipping in TMEM165-CDG. Clin Genet 87(1): 42-48. 10.1111/cge.12402
- Bammens R, Mehta N, Race V, Foulquier F, Jaeken J, Tiemeyer M, Steet R, Matthijs G, Flanagan-Steet H (2015). Abnormal cartilage development and altered N-glycosylation in Tmem165-deficient zebrafish mirrors the phenotypes associated with TMEM165-CDG. Glycobiology 25(6): 669-682. 10.1093/glycob/cwv009
- Shoemaker GK, Soya N, Palcic MM, Klassen JS (2008). Temperature-dependent cooperativity in donor-acceptor substrate binding to the human blood group glycosyltransferases. Glycobiology 18(8): 587-592. 10.1093/glycob/cwn043
- Gulberti S, Fournel-Gigleux S, Mulliert G, Aubry A, Netter P, Magdalou J, Ouzzine M (2003). The functional glycosyltransferase signature sequence of the human beta 1,3-glucuronosyltransferase is a XDD motif. J Biol Chem 278(34): 32219-32226. 10.1074/jbc.M207899200
- Boeggeman E, Qasba P-K (2002). Studies on the metal binding sites in the catalytic domain of β1,4-galactosyltransferase. Glycobiology 12(7): 395-407. 10.1093/glycob/cwf045
- Rayner JC, Munro S (1998). Identification of the MNN2 and MNN5 mannosyltransferases required for forming and extending the mannose branches of the outer chain mannans of Saccharomyces cerevisiae. J Biol Chem 273(41): 26836–26843. 10.1074/jbc.273.41.26836
- Romero PA, Herscovics A (1989). Glycoprotein biosynthesis in Saccharomyces cerevisiae: characterization of α-1,6-mannosyltransferase which initiates outer chain formation. J Biol Chem 264(4): 1946–1950. 2644248
- Striebeck A, Robinson DA, Schüttelkopf AW, van Aalten DM (2013). Yeast Mnn9 is both a priming glycosyltransferase and an allosteric activator of mannan biosynthesis. Open Biol 3(9): 130022. 10.1098/rsob.130022
- Wiggins CA, Munro S (1998). Activity of the yeast MNN1 α-1,3-mannosyltransferase requires a motif conserved in many other families of glycosyltransferases. Proc Natl Acad Sci U S A 95(14): 7945–7950. 10.1073/pnas.95.14.7945
- Antebi A, Fink GR (1992). The yeast Ca2+-ATPase homologue, PMR1, is required for normal Golgi function and localizes in a novel Golgi-like distribution. Mol Biol Cell 3(6): 633-654. 10.1091/mbc.3.6.633
- Vallee F, Lipari F, Yip P, Sleno B, Herscovics A, Howell PL (2000). Crystal structure of a class I alpha1,2-mannosidase involved in N-glycan processing and endoplasmic reticulum quality control. EMBO J 19(4): 581-588. 10.1093/emboj/19.4.581
- Chen JL, Ahluwalia JP, Stamnes M (2002). Selective effects of calcium chelators on anterograde and retrograde protein transport in the cell. J Biol Chem 277(38): 35682-35687. 10.1074/jbc.M204157200
- Snyder NA, Palmer MV, Reinhardt TA, Cunningham KW (2019). Milk biosynthesis requires the Golgi cation exchanger TMEM165. J Biol Chem 294(9):3181-3191. 10.1074/jbc.RA118.006270
- McEvoy J.P, Brudvig G.W (2006). Water-Splitting Chemistry of Photosystem II. Chem Rev 106(11):4455-4483. 10.1021/cr0204294
- Hoecker N, Leister D, Schneider A (2017). Plants contain small families of UPF0016 proteins including the PHOTOSYNTHESIS AFFECTED MUTANT71 transporter. Plant Signal Behav 12(2): e1278101. 10.1080/15592324.2016.1278101
- Lilley RM, Ralph PJ, Larkum AW (2010). The determination of activity of the enzyme Rubisco in cell extracts of the dinoflagellate alga Symbiodinium sp. by manganese chemiluminescence and its response to short-term thermal stress of the alga. Plant Cell Environ 33(6): 995-1004. 10.1111/j.1365-3040.2010.02121.x
- Papp-Wallace KM, Maguire ME (2006). Manganese Transport and the Role of Manganese in Virulence. Annu Rev Microbiol 60(1): 187-209. 10.1146/annurev.micro.60.080805.142149
- Juttukonda LJ, Skaar EP (2015). Manganese homeostasis and utilization in pathogenic bacteria. Mol Microbiol 97(2): 216-228. 10.1111/mmi.13034
- Colinet AS, Thines L, Deschamps A, Flémal G, Demaegd D, Morsomme P (2017). Acidic and uncharged polar residues in the consensus motifs of the yeast Ca2+ transporter Gdt1p are required for calcium transport. Cell Microbiol 19(7): e12729. 10.1111/cmi.12729
- Rosnoblet C, Legrand D, Demaegd D, Hacine-Gherbi H, de Bettignies G, Bammens R, Borrego C, Duvet S, Morsomme P, Matthijs G, Foulquier F (2013). Impact of disease-causing mutations on TMEM165 subcellular localization, a recently identified protein involved in CDG-II. Hum Mol Genet 22(14): 2914-2928. 10.1093/hmg/ddt146
- Lee JS, Kim MY, Park ER, Shen YN, Jeon JY, Cho EH, Park SH, Han CJ, Choi DW, Jang JJ, Suh KS, Hong J, Kim SB, Lee KH (2018). TMEM165, a Golgi transmembrane protein, is a novel marker for hepatocellular carcinoma and its depletion impairs invasion activity. Oncol Rep 40(3): 1297-1306. 10.3892/or.2018.6565
- Krzewinski-Recchi M-A, Potelle S, Mir A-M, Vicogne D, Dulary E, Duvet S, Morelle W, de Bettignies G, Foulquier F (2017). Evidence for splice transcript variants of TMEM165, a gene involved in CDG. Biochim Biophys Acta 1861(4): 737-748. 10.1016/j.bbagen.2017.01.011
–
ACKNOWLEDGMENTS
The work was supported by grant from the Fonds National de la Recherche Scientifique (FNRS, grant PDR-T.0206.16). L.T. was a research fellow at the “Fonds pour la Formation à la Recherche dans l’Industrie et dans l’Agriculture”.
COPYRIGHT
© 2020

From the Uncharacterized Protein Family 0016 to the GDT1 family: Molecular insights into a newly-characterized family of cation secondary transporters by Thines et al. is licensed under a Creative Commons Attribution 4.0 International License.








