
Reviews:
Microbial Cell, Vol. 8, No. 9, pp. 208 - 222; doi: 10.15698/mic2021.09.759
Understanding the pathogenesis of infectious diseases by single-cell RNA sequencing

1 Laboratory of Bacterial Pathogenesis, Department of Microbiology and Immunology, Institutes of Medical Sciences, Shanghai Jiao Tong University School of Medicine, Shanghai 200025, China.
2 Department of Infectious Diseases, Shanghai Ruijin Hospital, Shanghai 200025, China.
Keywords: single-cell RNA sequencing, infectious diseases, bacteria, viruses, fungi, parasites, immune response.
Received originally: 19/02/2021 Received in revised form: 13/07/2021
Accepted: 21/07/2021
Published: 04/08/2021
Correspondence:
Yu-Feng Yao, Rm 710, Bldg 5, 280 S. Chongqing Rd, Shanghai, China 200025; Tel: 021-63846590-776523; Mobile: 13817347153; Fax: 021-64671226; yfyao@sjtu.edu.cn
Conflict of interest statement: The authors have declared no conflict of interest.
Please cite this article as: Wanqiu Huang, Danni Wang and Yu-Feng Yao (2021). Understanding the pathogenesis of infectious diseases by single-cell RNA sequencing. Microbial Cell 8(9): 208-222. doi: 10.15698/mic2021.09.759
Abstract
Infections are highly orchestrated and dynamic processes, which involve both pathogen and host. Transcriptional profiling at the single-cell level enables the analysis of cell diversity, heterogeneity of the immune response, and detailed molecular mechanisms underlying infectious diseases caused by bacteria, viruses, fungi, and parasites. Herein, we highlight recent remarkable advances in single-cell RNA sequencing (scRNA-seq) technologies and their applications in the investigation of host-pathogen interactions, current challenges and potential prospects for disease treatment are discussed as well. We propose that with the aid of scRNA-seq, the mechanism of infectious diseases will be further revealed thus inspiring the development of novel interventions and therapies.
INTRODUCTION
Infectious diseases have always been serious threats to public health. Excellent examples are the recent global epidemic of Coronavirus disease 2019 (COVID-19), long-standing influenza, HIV, Salmonella, as well as other bacterial infections. To clarify the pathogenesis of these diseases, it is necessary to understand the interactions between the host and the pathogen. Although traditional phenotypic measurements and bulk transcript analysis can provide some insights into pathogenesis, neither the heterogeneity of individual cell populations nor the pathogenic states can be appreciated. With the development of single-cell RNA sequencing (scRNA-seq), a new technique, which measures the transcriptome at the single-cell level, it is possible to study cell behaviors at a higher resolution. The use of scRNA-seq has refined the human cell landscape [1] and driven progress in various areas including immunology [2], developmental biology [3], oncology [4], and infectious diseases. Single cell approaches allow for identification of cellular heterogeneity during infections as well as subpopulation differences that influence outcomes and individual differences. Recent advances in scRNA-seq analysis of infections have provided for a new understanding of host-pathogen interactions. In this review, we briefly introduce technological improvements in scRNA-seq, highlight reports that interrogate microbial pathogenicity and host immune responsiveness at the single-cell level, describe the current limitations of scRNA-seq, and discuss exciting prospects for the study of infections and possible clinical application.
THE TECHNOLOGY ADVANCES IN SINGLE-CELL RNA SEQUENCING
The first transcriptome analysis at the single-cell level was conducted by Tang et al. in 2009 [5]. Since then, this technology has been continuously improved to meet different needs, leading to the emergence of several novel methods such as: SMART-seq2 [6], Drop-seq [7], inDrop [8], CEL-seq2 [9][10], and MARS-seq [11]. To date, scRNA-seq has developed into a mature workflow, including single cell isolation, cell lysis, conversion of RNA into cDNA with amplification, library construction, sequencing, and analysis of the high-throughput data. These new technologies were developed by improving key steps including cell separation, library construction, sequencing depth, and quality.
–
The emergence of the use of barcodes [12] and unique molecular identifiers (UMI) was a huge advance [13]. The single-cell tagged reverse transcription (STRT) sequencing, which first introduced cell-specific barcoding at the reverse transcription stage, enabled highly multiplexed analysis [12]. After that, the addition of UMIs identified each molecule in a population as distinct, as a random DNA sequence label or an aliquot of a complex mixture [14]. Multiple scRNA-seq methods such as CEL-seq, Drop-seq, and MARS-seq assess the combination of barcodes and UMIs, providing for high throughput and sensitivity. However, multiplexing cDNA amplification sacrifices full-length coverage. These methods profile only the 5′- or 3′-terminus of the transcripts. In contrast, SMART-seq2 does not use barcodes or UMIs. The cDNA libraries are generated from individual cells, providing full-length transcripts [6] that increase scalability and availability. A newly developed multiple annealing and dC-tailing-based quantitative single cell RNA sequencing (MATQ-seq) not only captures the full-length RNA and genuine biological variation between whole transcriptomes [15] but also adds UMIs reducing bias with higher sensitivity and lower technical noise.
–
Another improvement worth mentioning is the application of maturing sequencing platforms. Previous methods, e.g., CEL-seq [9], which was inefficient and error-prone, were mainly plate-based. CEL-seq2 [10] employs an automated microfluidic platform from Fluidigm (C1 platform). With MARS-seq, a high-throughput implementation of the original CEL-seq method [11], cells are sorted by fluorescence-activated cell sorting (FACS). The newly developed Drop-seq [7] and inDrop [8] use nanoliter droplets to capture single cells. For Microwell-seq, a high-throughput and low-cost platform, individual cells are trapped in an agarose microarray and mRNAs are captured with magnetic beads [16]. All these innovative platforms have improved cell sorting accuracy. The availability of commercial platforms such as the Chromium system from 10×Genomics improves scRNA-seq efficiency by automation and lowers cost as well.
–
Briefly, even though various technologies have been developed, it is necessary to carefully consider the most suitable method for analysis based on actual situations and experimental purposes. A comparative analysis of prominent scRNA-seq methods revealed that Drop-seq is more cost-efficient when quantifying the transcriptomes of large numbers of cells at low sequencing depth. Single cell RNA barcoding and sequencing (SCRB-seq), with massively parallel single-cell RNA sequencing (MARS-seq), is preferable when quantifying transcriptomes of fewer cells [17].
BACTERIAL INFECTION
The outcomes of an infection are complicated interactions of the pathogen and the host involving multiple biological factors. Pathogen virulence and growth state, host immunity, diverse cell types, and tissue microenvironments all impact disease progression and antimicrobial treatment. ScRNA-seq has become a powerful tool to probe cell-to-cell variability and uncover both host and bacterial factors that influence the severity of infection. To date, many scRNA-Seq studies have been performed to investigate the host-pathogen interactions (Table 1).
–
| Table 1. The applications of scRNA-seq in infection. |
–
One well-studied example of infectious diseases is Salmonella infection, a common food-borne pathogen that can produce acute or chronic symptoms, either a limited gastroenteritis or a systemic infection [18]. Recent studies have shown that the intracellular bacterial heterogeneity is influenced by host cell microenvironments which, in turn, can produce differential host cell immune responses. During the infection, macrophages are favorable niches for Salmonella survival and proliferation. The scRNA-seq expression profiling of infected macrophages revealed that the induction of a macrophage type I interferon (IFN) response correlated with variable PhoPQ activity of invading Salmonella[19]. When macrophages came across a subset of bacteria with highly modified lipopolysaccharides by PhoPQ, the macrophages tended to have a high type I IFN response. Meanwhile, macrophages harboring non-growing Salmonella displayed hallmarks of the proinflammatory M1 polarization state that differed little from bystander cells [20]. The non-growing bacteria did not trigger additional immune recognition by intracellular receptors. However, intracellular growing bacteria induced an M2-like anti-inflammatory response in macrophages, indicating that intracellular Salmonella were capable of escaping from the host defense by reprogramming macrophage polarization [20]. These data suggest that gene expression heterogeneity among infected cells creates diverse environments for Salmonella to either persist or exploit its host. In addition, the bacterial pathogenicity also plays an important role in regulating the host cell state and immune response.
–
Furthermore, different S. Typhimurium strains have been shown to produce marked differences during infection [21]. Compared with non-invasive Salmonella, the highly invasive and multi-drug resistant S. Typhimurium strain ST313 could produce a heterogeneous innate immune response that exploited divergent evasion strategies for dissemination in vivo. MoDCs infected with invasive Salmonella differentially regulated genes associated with endosomal trafficking and antigen presentation pathways. Invasive Salmonella induced higher expression of IL10 and MARCH1 but lower expression of CD83, allowing evasion of adaptive immune supervision.
–
Moreover, investigations have restricted analysis to only eukaryotic transcripts, thereby losing the ability to decipher the heterogeneity of both host and bacteria at the same time. The development of scDual-Seq enables the capture of host and pathogen transcriptomes simultaneously at the single cell level [22]. By utilizing this method to study the process of individual macrophages, the authors showed the rate of S. Typhimurium infection was non-uniform, supportive by the evidence showing the co-existence of all three cell subpopulations. These three cell states also showed evidence for a linear progression through consecutive stages of infection. However, the experiments were limited in that a high multiplicity of infection (MOI) could cause a variable number of bacteria in infected macrophages, thus masking certain infectious stages and phenotypes.
–
Another suitable model for studying host-bacterial interaction is Mycobacterium tuberculosis (Mtb), which sickens millions of people with tuberculosis (TB). Lungs infected with Mtb contain several coexisting lesion types such as solid cellular granulomas of densely packed macrophages and necrotic granulomas with an outer ring of T and B lymphocytes [23]. Technological advances in scRNA-seq helped to reveal the heterogeneity between different cell types and subpopulations during Mtb infection [24]. Gene expression shifts associated with cell growth and metabolism were found among cell clusters in response to Mtb. The complex dynamic of host and bacteria can result in distinct symptoms from latent TB infection (LTBI), in which patients remain clinically asymptomatic, to active TB, a contagious state in which patients may suffer from cough, fever and night sweats [25]. A small proportion of LBTI patients undergo progression into active TB. For better TB control, it is necessary to identify these populations by monitoring reliable biomarkers. Since the intensity of immune response and alterations in immune cell composition are critical indicators of severity of disease, many efforts have been made to investigated the transcript profiles of immune cells in peripheral blood and lesions. A recent scRNA-seq study compared the transcriptomes of peripheral blood mononuclear cells (PBMC) from healthy controls (HC), (LTBI and active TB patients. The results showed that there was a gradual depletion of the cytotoxic natural killer (NK) cell subset (CD3−CD7+GZMB+) during these three states [26]. The subset frequency also increased after anti-TB treatment, which confirmed that the frequency change in NK cells was involved in host disease severity and could be used as a novel biomarker to discriminate patients with TB from LTBI and HC.
–
The immune response to infectious agents is an orchestrated process that involves numerous cell types and pathways. Ronnie et al. comprehensively characterized the initial 48 h of the innate immune response to diverse pathogens by MARS-seq [27]. They found that most lymph node cell types showed little pathogen specificity and that antigen-specific immunity was driven by antigen-carrying dendritic cells and monocytes. The NK-driven IFN-γ response initiated a monocyte-specific signaling cascade that promoted Th1 development. Taken together, these data demonstrate an innate immune heterogeneity in response to a wide range of pathogens. This knowledge may provide insight into the development of safe and effective vaccines.
VIRAL INFECTION
Viral infections are always of urgent public concern because of their high degree of transmissibility and pathogenic severity. Representative examples are influenza, HIV, and SARS-CoV. Advances in single cell technology coupled with mathematical modeling and computer science provide new horizons for understanding virus and host interactions.
–
COVID-19, a newly emerged severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has become an ongoing global health emergency. Upon entry into the host cell, SARS-CoV-2 can replicate quickly and trigger a strong immune response leading to an acute respiratory syndrome, pulmonary tissue damage, and multiple organ failure [28]. This highly contagious disease progresses rapidly and so far has caused tens of millions of cases. Therefore, it is an urgent need to determine the pathogenic basis of this disease in order to improve existing prevention and treatment strategies. Single cell technology has shown its unique advantages in relevant investigations that uncovered sensitive cell types and the heterogeneity of the host immune response, each of which provided valuable insights into vaccine development.
–
To invade cells, SARS-CoV-2 binds host angiotensin-converting enzyme 2 (ACE2) with its spike (S) protein and utilizes a type II transmembrane serine protease 2 (TMPRSS2) for priming and activation of the protein [29][30]. Using scRNA-seq expression analysis of ACE2 and TMPRSS2, different cell types, tissues, and organs were assessed. The lung, kidney, bladder, and ileum were found to have the greatest expression [31]. Within these organs, oral mucosa [32], nasal epithelial cells [33], type II alveolar cells [31], and bronchial transient secretory cells [34] expressed the highest levels of ACE2 and TMPRSS2. As such, these cells have increased sensitivity to SARS-CoV-2 infection. Based on evidence obtained from a single cell RNA expression map of human coronavirus entry factors, spermatogonia and prostate endocrine cells were permissive to SARS-CoV-2 infection [34], suggesting that males were more vulnerable to coronavirus infection [35]. Multiple risk factors like age, gender and cigarette smoking contribute to the infection progression. The chronic smoke exposure increases lung ACE2 expression and triggers the expansion of ACE2+ secretory cells in the respiratory tract [35]. Aging increases the gene expression associated with SARS-CoV-2 susceptibility and COVID-19 can promote age-induced immune cell polarization [36]. The expression level of TMPRSS2 is increased in older adults compare to children, which could be a reason for the higher risk of severe disease among the aged [37]. Furthermore, several studies demonstrated ACE2 to be an IFN-stimulated gene in humans that is upregulated by viral infection [38]. These data allow for the construction of a risk map of vulnerable cell types, revealing underlying virus transmission mechanisms (Figure 1).
–
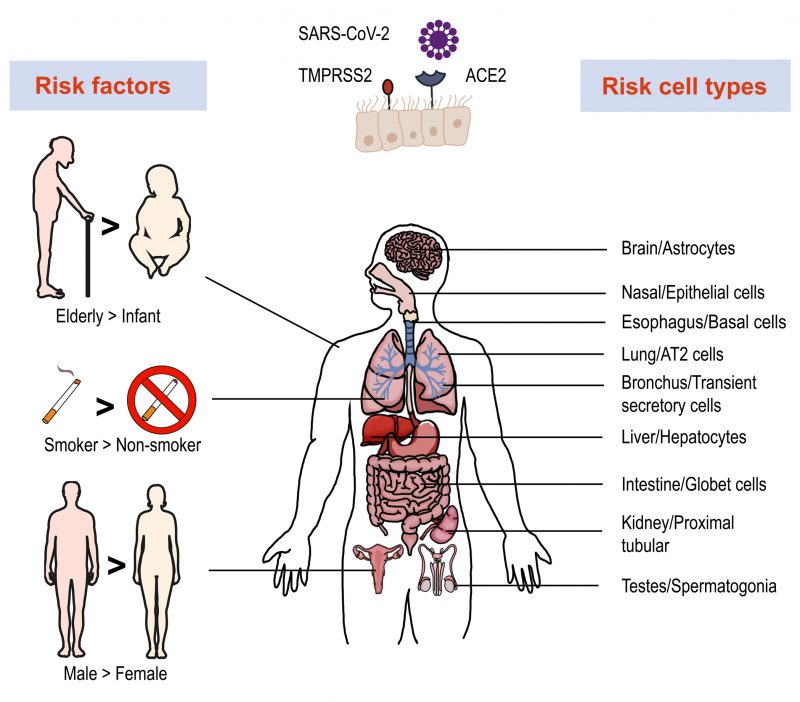 | FIGURE 1: Risk factors, vulnerable organs and cell types involved in SARS-Cov-2 infection. Single-cell RNA-seq revealed high expression of ACE2 and TMPRSS2 in multiple organs and cell types, which are more vulnerable to SARS-Cov-2 infection. Several factors play important roles in infection and disease process, such as age, smoking and gender. |
–
COVID-19 patients present a wide spectrum of clinical manifestations, which range from asymptomatic infection to severe pneumonia [39]. The relationship between disease severity and the host immune response is not fully understood. Single cell analysis of immune cells and local landscape signature provide for an understanding of the dynamic immune response, which has great heterogeneity at the cellular level and differences among individuals.
–
A newly developed computational framework, Viral-Track, enables the global scanning of unmapped viral RNA in scRNA-seq data. Integrating these data with the host transcriptome distinguishes infected cells from bystander cells and manifests specific virus-induced expression [40]. This method was applied to bronchoalveolar-lavage (BAL) samples from severe and mild COVID-19 patients and revealed the dramatic diversity of immune cell compartments and responses. For example, the myeloid compartment in mild patients was enriched in alveolar macrophage and plasmacytoid dendritic cells (pDCs). In severe patients, accumulated myeloid cells were neutrophils, FCN+ monocytes, and monocyte-derived macrophages, which expressed a viral hallmark IFN type I response [41] and upregulation of inflammatory chemokine genes, e.g., CCL18. Similarly, severe COVID-19 infections were shown to be associated with developing neutrophils [42][43] and abundant proinflammatory monocyte-derived macrophages [44] with an imbalance of the T cell compartment and T cell function. Proliferating cytotoxic CD8+ T cells and CD4+ follicular helper T cells (Tfh) were common in COVID-19 patients with reduced proportions of regulatory T cells (Treg) [45]. In moderate cases of COVID-19, CD8+ T cells were markedly expanded with higher levels of effector molecules, e.g., XCL1 [44], and a naïve phenotype for CD4+ T cells in severe cases [40].
–
In addition to the virus-stimulated alterations in cell composition, specific host immune response pathways provide insight into pathogenesis. SARS-CoV-2 infection induced the activation of the STAT1/IRF3 pathway and increased the expression levels of IL6R (interleukin 6 receptor) and IL6ST (interleukin 6 signal transducer), which may have a synergistic effect with elevated IL-6 to induce a strong inflammatory response [46]. These results provide evidence that cytokine inhibition may be a reliable strategy to attenuate severe inflammatory responses in COVID-19 patients, e.g., disruption of IL-6 and IL-6R binding [47], CCR1 and/or CCR5 pathway [48]. Additionally, host factors and pathways involved in key elements of the SARS-CoV-2 viral life cycle have been shown to have a role in regulating the infection [49]. For example, the ATPase proton pump which interacts with SARS-CoV-2 non-structural protein 6 (nsp6) and RAB7A which interacts strongly with non-structural protein 7 (nsp7). The loss of RAB7A reduced viral entry by sequestering ACE2 receptors via altered endosomal trafficking. The upregulation of cholesterol biosynthesis pathway with the small molecule amlodipine led to reduction of viral infection. It is possible that changes in lipid composition directly impacted SARS-CoV-2 virion maturation and infectivity, indicating potential new therapeutic targets [49].
–
Now that many patients have recovered from the virus infection, studying the immune system of these people will provide valuable information for prevention and treatments. Neutralizing antibodies are a powerful weapon to block virus entry. Through analyzing single B cell RNA/VDJ data of COVID-19 convalescent patients, novel potent neutralizing antibodies and S protein-binding antibodies have been identified, among which BD-368-2 showed a great therapeutic effect and GD1-69 showed a powerful neutralizing activity [50][51]. Multiple investigations have demonstrated antibody cocktails to largely prevent viral mutation escape which produces an optimal antiviral effect [52]. ScRNA-seq technology enables the discovery of new neutralizing antibodies, accelerating development of new antiviral drugs and vaccines.
–
Individual cellular immunity participating in virus-host interaction is crucial for controlling virus elimination. A recent single cell transcriptome analysis revealed a dynamic cellular program during longitude HIV acute infection, such as NK cell expansion, naïve CD4+ T cell differentiation, and a rapid rise in plasma viremia coupled with cell-type specific interferon-stimulated gene (ISG) upregulation [53]. These cell type frequency shifts at different time points may shed light on how the immune response were orchestrated according to the infection progression. In accordance with changes in cellular phenotype, the release of pro-inflammatory factors also showed a time-course fluctuation [54]. During the early stage (day 1-3 post infection) of influenza A virus (IAV) infection, a group of PD-L+ neutrophils was the major contributor to the first wave of pro-inflammatory factors including Ccl3, Cxcl10, TNF-α, and IL1α. High level of virus HA mRNA and protein were also detected in the cell population. The second wave was mainly generated by another subset of Pf4+-macrophages at a later stage (day 7 post infection), which may be the precursor of alveolar macrophages. Enhancing the immune response against pathogens and reducing the cytokine storm are the major therapeutic aims of infectious disease. Analysis of these dynamic changes at single cell level could give deep insights into a more complete understanding of immunopathogenesis and offer possible targets for clinical treatment.
PARASITIC INFECTION
Parasitic infection is an important worldwide cause of human disease such as Chagas disease, African trypanosomiasis, amebiasis, leishmaniasis, ascariasis, and schistosomiasis, leading to millions of deaths [55]. Malaria is a prevalent parasitic disease with 229 million victims worldwide in 2019 as reported by the World Health Organization (WHO). The disease is caused by a unicellular eukaryotic parasite, Plasmodium spp., that is transmitted to humans by Anopheles spp. mosquitoes [56]. Plasmodium falciparum is the most virulent cause of malaria. During human infection, the parasite attacks the liver and then invades the vasculature where red blood cells are infected resulting in an intra-erythrocytic development stage [57]. During this blood stage, the fast asexual-replication of parasites produces the clinical manifestation of the disease. The parasite then converts to a non-replicating sexual gametocyte form, which can be transmitted to female mosquito during a blood meal [58]. Understanding the complex life cycle of Plasmodium is fundamental for pathogen elimination and disease treatment. Although these distinct parasitic stages have been investigated at the overall population level [57], little is known about the variation between individual parasites.
–
ScRNA-seq promises a precise examination of transcriptional expression heterogeneity during parasitic stage switching and immune response. The transcriptional activation of AP2-G is known as the master regulator to initialize the sexual commitment [59]. AP2-G+ mature schizonts specifically upregulated epigenetic regulators like histone-modifying enzymes, which facilitated the subsequent gametocyte development [60]. Individual parasite transcripts showed distinct stage-specific transcriptional transitions and revealed differential routes P. falciparum converted into gametocytes [61][62][63]. A set of sex-specific genes involved in sequestrating gametocytes were found to function during immune evasion as well as transmission from human to mosquitos. Meanwhile, coupled with the synchronized single-cell transcriptome of infected red blood cells, novel gene signature that could be used as an indication to discriminate between sexual and asexual stages were identified [64]. In brief, scRNA-seq has allowed for high-resolution mapping of the life cycle of Plasmodium spp., providing a fundamental resource for the investigation of parasite biology and the study of malaria pathogenesis [65].
–
During the blood-stage of Plasmodium infection, CD4+ T cells and myeloid cells both play important roles in controlling the progress of disease, in which the intercellular communication fosters the immune regulation [66]. The CD4+ T-effector cell TCRβ repertoires undergo a polyclonal expansion dominated by TRBV3 gene usage in the acute phase of P. chabaudi infection [67]. Two types of CD4+ T cells are the main sources of protecting against malaria: (1) T helper (Th1) cells that secret IFN-γ and stimulate phagocytic cells to capture and kill parasites [68], and (2) Tfh cells that promote development of antigen-specific B cells that produce anti-parasitic antibodies [69]. During the activation and differentiation of CD4+ T cells, myeloid cells were shown to play a vital regulatory role, not only the dendritic cells presented the antigen stimulation, but inflammatory monocytes could also support a Th1 fate at the Th1/Tfh bifurcation [70]. Furthermore, CD4+ T cell-derived macrophage stimulating factor (MCSF) facilitated the expansion and activation of specific myeloid subsets such as macrophages during P. chabaudi infection, which contributed to parasite clearance and infection control [71]. Heterogeneity revealed by scRNA-seq provides insights into the key molecular and immunological mechanisms of parasite infection as well as the identification of potential targets for parasite elimination and vaccine development [72].
FUNGAL INFECTION
Fungal disease is also a threatening heath problem, causing over 1.5 million deaths worldwide each year [73]. With advances in medical and surgical therapy of many different diseases, opportunistic fungi have become a major source of nosocomial infections especially among immunocompromised patients [74]. Candidiasis and aspergillosis account for a large proportion of these infections [75]. Candidiasis is caused by Candida, an opportunistic fungus that colonizes the skin and mucosa, which can lead to bloodstream infections, known as candidemia [76]. Single cell transcriptomic analysis showed that Candida stimulation induced dramatic and differential gene expression within CD4+ T cells, NK cells, and monocytes, while the upregulation of the IFN I pathway was consistent across all cell types [77]. Combination with published bulk RNA-seq data revealed that Candida-response expression quantitative trait loci (eQTL) was associated with disease susceptibility. Of note, LY86 was found to be a potential candidemia-risk allele, with reduction of LY86 weakening the migration of monocytes, thus increasing susceptibility for candidemia [77].
–
Nevertheless, the use of scRNA-seq to determine the transcriptomes of pathogens and hosts during fungal infection are still in their infancy. Most investigations employ single cell transcriptomics that focus on life cycles, cell states, taxonomic diversities, and ecological interactions [78][79], rather than fungal infection. Given the increasing morbidity and mortality of fungal disease, deeper and more detailed explorations of fungal-host interactions are urgently needed.
CHALLENGES AND LIMITATIONS
Although single cell technology has revolutionized many fields, there are still several challenges impeding deep exploration. A major limiting factor is the amount of available nucleic acid in that one mammalian cell contains only 10 pg of total RNA [80]. This limit may result in artificial zeros either systematically or accidently, thus hindering complete downstream analysis [81]. Using computational statistical models, it may be possible to circumvent this limitation by modulation of sample variation and noise by use of VIPER [82] and scSDAEs [83]. With regard to infection, most studies have focused on either the host or the pathogen, whereas dual RNA-seq allows analysis of the transcripts of both host and bacteria simultaneously [84], but not at the single-cell bacterial level. However, improved scDual RNA-seq does enable profiling of both host and bacterial transcripts in one individually infected mammalian cell [22]. It is important to note that the detection of intracellular bacteria is technically difficult since a single bacterium contains only 100 fg of total RNA. Further, the mRNA of bacteria and some virus like hepatitis C virus and dengue virus lack poly (A) tails which are required for library construction. Enzymatic addition of poly (A) tails, adaptor ligation, and/or random priming for amplification [22][85] address this problem but also require additional steps to deplete redundant rRNA [86]. Advanced methodologies for characterization of single infected cells of both host and pathogen are needed.
–
Infection is a complicated process, with outcomes of infection dependent on the heterogeneity of host and pathogen. Host heterogeneity is at the cell type level, in other words, the vulnerability of host cells and their response to pathogens are attributed to the host cell type [20]. Heterogeneity expressed by different pathogens results in various host cell responses, resulting in local or systemic infection. The biological implications of these results are worth exploration with spatial and trajectory information of great value for analyzing the dynamics of host-pathogen interactions. To obtain single cell transcriptomes, traditional isolation of single cells usually digests whole tissues or captures the entire cell [87], failing to preserve spatial context. Recently, pioneering work combining single molecule fluorescence in situ hybridization (smFISH) with RNA quantification was used to interrogate spatial gene expression at the single cell level [88]. Subsequently, efforts have been made to increase the number of transcripts measured within a single cell by use of MERFISH [89] and seqFISH+ [90]. Integrating microarray-based spatial transcriptomics can assist in mapping the architecture of scRNA-seq-defined subpopulations [91]. Related computational analysis strategies such as trendsceek [92], which measures single-cell spatial gene expression have been developed as well.
–
Both infection and immune response are continuous processes while most scRNA-seq samples are taken at discrete time periods. One mean by which to supervise this dynamic process is to implement real-time monitoring, which is technically challenging and expensive. Another solution is the use of computer models to simulate potential cell developmental paths based on similarities in expression patterns, which are referred to as trajectory inference or pseudo-time analysis [93]. Recently, utilizing kinetic scRNA-seq, Abbas et al. characterized pDCs activation trajectories during mouse cytomegalovirus (MCMV) infection by pseudo-time analysis [94]. They found pDCs to manifest multiple functions (IFN-I production and T cell activation) in a time and space tightly orchestrated manner. Likewise, Khatun et al. evaluated CD4+ T cell clonal expansion and differentiation trajectories in response to acute lymphocytic choriomeningitis virus (LCMV) infection [95]. Although a number of analysis technologies and methodologies are available, method consideration is mainly dependent on dataset dimension and trajectory topology [96]. These innovative tools will enable the recall analysis of in vivo infections, which will provide for detailed identification of pathogenic mechanism, even though limitations still exist.
PERSPECTIVES
Advances in the application of scRNA-seq for the study of infections have dramatically broadened our understanding of the molecular details of host-pathogen interactions (Figure 2). These advances have allowed for the prediction of infection outcomes and the discovery of new diagnostic biomarkers and therapeutic targets [97]. For example, Ben-Moshe et al. developed a deconvolution algorithm for predicting bacterial infection outcomes based on the scRNA-seq data of human PBMCs infected with Salmonella[98]. Applying this algorithm to bulk RNA-seq data from cohorts of TB patients during different stages of disease, they not only distinguished active TB patients from healthy individuals but also identified individuals with higher risk for development of active TB. Considering the urgent need for new drugs and vaccines for COVID-19, Alakwaa et al. developed a bioinformatic pipeline to prioritize drug candidates based on existing scRNA-seq data and drug perturbation databases, which identified four drugs including didanosine with potential efficacy [99]. Such studies offer advanced treatment options for infectious disease therapy and intervention.
–
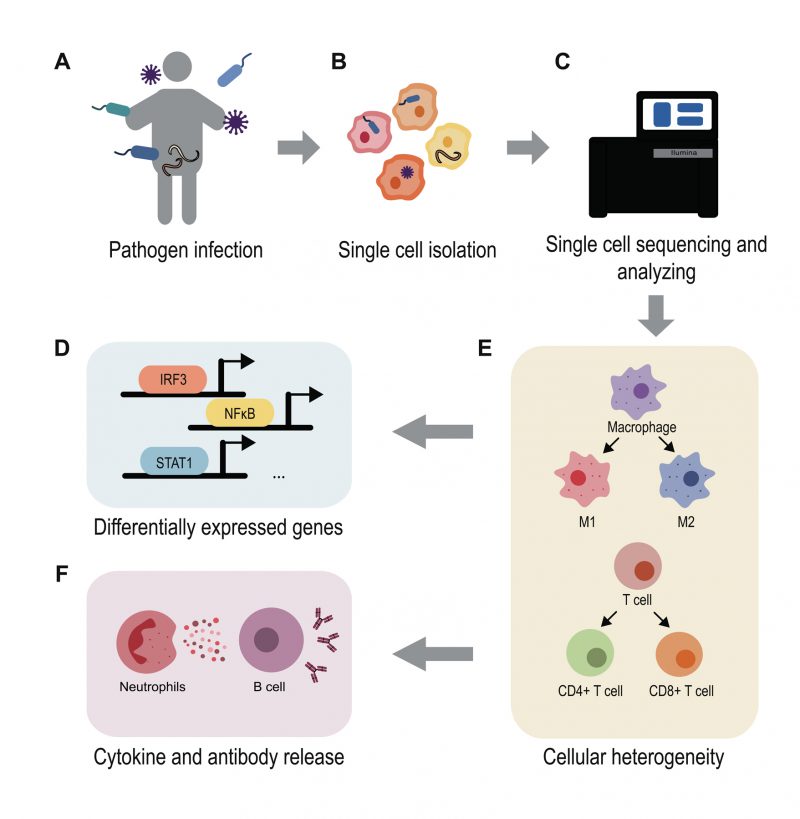 | FIGURE 2: Using single-cell RNA-seq to understand host-pathogen interactions. Pathogens like bacteria, viruses, fungi and parasites can infect hosts and cause various infectious diseases (A). Using scRNA-seq methods enable the study of host and pathogen interactions at a high-resolution level (B-C). The single cell transcriptomes reveal the differentially gene expression map during infection (D), demonstrate the cell heterogeneity and immune cell development (E), uncover the mechanism behind the inflammatory and anti-microbial response (F). The application of scRNA-seq highly contributes to infection research. |
–
In addition to the spatial transcriptome, which reveals spatial cellular information, emerging multiple-omics single cell analysis can link chromatin and protein features to gene expression in a single cell [100], e.g., scDam and T-seq [101] and SHARE-seq [102]. Combinatorial approaches will further shed light on the multidimensional features of a single cell, refining characterization of the cell heterogeneity during infection. Overall, advances in scRNA-seq technology have greatly improved our understanding of host-pathogen interaction and made fundamental contributions to the development of new strategies for the control of infectious disease [103]. These advances have enabled deeper understanding of the infectious progress, although analyses of these types have just been initiated. Further efforts are still required and with the continuous creation of new approaches, the mechanism of the infectious diseases will gradually be uncovered, as well as new drugs and effective treatments.
REFERENCES
- Han X, Zhou Z, Fei L, Sun H, Wang R, Chen Y, Chen H, Wang J, Tang H, Ge W, Zhou Y, Ye F, Jiang M, Wu J, Xiao Y, Jia X, Zhang T, Ma X, Zhang Q, Bai X, Lai S, Yu C, Zhu L, Lin R, Gao Y, Wang M, Wu Y, Zhang J, Zhan R, Zhu S, et al. (2020). Construction of a human cell landscape at single-cell level. Nature 581(7808): 303-309. 10.1038/s41586-020-2157-4
- Stubbington MJT, Rozenblatt-Rosen O, Regev A, Teichmann SA (2017). Single-cell transcriptomics to explore the immune system in health and disease. Science 358(6359): 58-63. 10.1126/science.aan6828
- Su T, Stanley G, Sinha R, D’Amato G, Das S, Rhee S, Chang AH, Poduri A, Raftrey B, Dinh TT, Roper WA, Li G, Quinn KE, Caron KM, Wu S, Miquerol L, Butcher EC, Weissman I, Quake S, Red-Horse K (2018). Single-cell analysis of early progenitor cells that build coronary arteries. Nature 559(7714): 356-362. 10.1038/s41586-018-0288-7
- Jackson HW, Fischer JR, Zanotelli VRT, Ali HR, Mechera R, Soysal SD, Moch H, Muenst S, Varga Z, Weber WP, Bodenmiller B (2020). The single-cell pathology landscape of breast cancer. Nature 578(7796): 615-620. 10.1038/s41586-019-1876-x
- Tang F, Barbacioru C, Wang Y, Nordman E, Lee C, Xu N, Wang X, Bodeau J, Tuch BB, Siddiqui A, Lao K, Surani MA (2009). mRNA-Seq whole-transcriptome analysis of a single cell. Nat Methods 6(5): 377-382. 10.1038/nmeth.1315
- Picelli S, Björklund ÅK, Faridani OR, Sagasser S, Winberg G, Sandberg R (2013). Smart-seq2 for sensitive full-length transcriptome profiling in single cells. Nat Methods 10(11): 1096-1098. 10.1038/nmeth.2639
- Macosko Evan Z, Basu A, Satija R, Nemesh J, Shekhar K, Goldman M, Tirosh I, Bialas Allison R, Kamitaki N, Martersteck Emily M, Trombetta John J, Weitz David A, Sanes Joshua R, Shalek Alex K, Regev A, McCarroll Steven A (2015). Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 161(5): 1202-1214. 10.1016/j.cell.2015.05.002
- Klein Allon M, Mazutis L, Akartuna I, Tallapragada N, Veres A, Li V, Peshkin L, Weitz David A, Kirschner Marc W (2015). Droplet Barcoding for Single-Cell Transcriptomics Applied to Embryonic Stem Cells. Cell 161(5): 1187-1201. 10.1016/j.cell.2015.04.044
- Hashimshony T, Wagner F, Sher N, Yanai I (2012). CEL-Seq: Single-Cell RNA-Seq by Multiplexed Linear Amplification. Cell Rep 2(3): 666-673. 10.1016/j.celrep.2012.08.003
- Hashimshony T, Senderovich N, Avital G, Klochendler A, de Leeuw Y, Anavy L, Gennert D, Li S, Livak KJ, Rozenblatt-Rosen O, Dor Y, Regev A, Yanai I (2016). CEL-Seq2: sensitive highly-multiplexed single-cell RNA-Seq. Genome Biol 17: 77. 10.1186/s13059-016-0938-8
- Jaitin DA, Kenigsberg E, Keren-Shaul H, Elefant N, Paul F, Zaretsky I, Mildner A, Cohen N, Jung S, Tanay A, Amit I (2014). Massively Parallel Single-Cell RNA-Seq for Marker-Free Decomposition of Tissues into Cell Types. Science 343(6172): 776. 10.1126/science.1247651
- Islam S, Kjällquist U, Moliner A, Zajac P, Fan JB, Lönnerberg P, Linnarsson S (2011). Characterization of the single-cell transcriptional landscape by highly multiplex RNA-seq. Genome Res 21(7): 1160-1167. 10.1101/gr.110882.110
- Kivioja T, Vähärautio A, Karlsson K, Bonke M, Enge M, Linnarsson S, Taipale J (2012). Counting absolute numbers of molecules using unique molecular identifiers. Nat Methods 9(1): 72-74. 10.1038/nmeth.1778
- Islam S, Zeisel A, Joost S, La Manno G, Zajac P, Kasper M, Lönnerberg P, Linnarsson S (2014). Quantitative single-cell RNA-seq with unique molecular identifiers. Nat Methods 11(2): 163-166. 10.1038/nmeth.2772
- Sheng K, Cao W, Niu Y, Deng Q, Zong C (2017). Effective detection of variation in single-cell transcriptomes using MATQ-seq. Nat Methods 14(3): 267-270. 10.1038/nmeth.4145
- Han X, Wang R, Zhou Y, Fei L, Sun H, Lai S, Saadatpour A, Zhou Z, Chen H, Ye F, Huang D, Xu Y, Huang W, Jiang M, Jiang X, Mao J, Chen Y, Lu C, Xie J, Fang Q, Wang Y, Yue R, Li T, Huang H, Orkin SH, Yuan GC, Chen M, Guo G (2018). Mapping the Mouse Cell Atlas by Microwell-Seq. Cell 172(5): 1091-1107.e1017. 10.1016/j.cell.2018.02.001
- Ziegenhain C, Vieth B, Parekh S, Reinius B, Guillaumet-Adkins A, Smets M, Leonhardt H, Heyn H, Hellmann I, Enard W (2017). Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol Cell 65(4): 631-643.e634. 10.1016/j.molcel.2017.01.023
- Kurtz JR, Goggins JA, McLachlan JB (2017). Salmonella infection: Interplay between the bacteria and host immune system. Immunol Lett 190: 42-50. 10.1016/j.imlet.2017.07.006
- Avraham R, Haseley N, Brown D, Penaranda C, Jijon HB, Trombetta JJ, Satija R, Shalek AK, Xavier RJ, Regev A, Hung DT (2015). Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses. Cell 162(6): 1309-1321. 10.1016/j.cell.2015.08.027
- Saliba AE, Li L, Westermann AJ, Appenzeller S, Stapels DA, Schulte LN, Helaine S, Vogel J (2016). Single-cell RNA-seq ties macrophage polarization to growth rate of intracellular Salmonella. Nat Microbiol 2: 16206. 10.1038/nmicrobiol.2016.206
- Aulicino A, Rue-Albrecht KC, Preciado-Llanes L, Napolitani G, Ashley N, Cribbs A, Koth J, Lagerholm BC, Ambrose T, Gordon MA, Sims D, Simmons A (2018). Invasive Salmonella exploits divergent immune evasion strategies in infected and bystander dendritic cell subsets. Nat Commun 9(1): 4883. 10.1038/s41467-018-07329-0
- Avital G, Avraham R, Fan A, Hashimshony T, Hung DT, Yanai I (2017). scDual-Seq: mapping the gene regulatory program of Salmonella infection by host and pathogen single-cell RNA-sequencing. Genome Biol 18(1): 200. 10.1186/s13059-017-1340-x
- Furin J, Cox H, Pai M (2019). Tuberculosis. Lancet 393(10181): 1642-1656. 10.1016/s0140-6736(19)30308-3
- Gierahn TM, Wadsworth MH, 2nd, Hughes TK, Bryson BD, Butler A, Satija R, Fortune S, Love JC, Shalek AK (2017). Seq-Well: portable, low-cost RNA sequencing of single cells at high throughput. Nat Methods 14(4): 395-398. 10.1038/nmeth.4179
- Pai M, Behr MA, Dowdy D, Dheda K, Divangahi M, Boehme CC, Ginsberg A, Swaminathan S, Spigelman M, Getahun H, Menzies D, Raviglione M (2016). Tuberculosis. Nat Rev Dis Primers 2: 16076. 10.1038/nrdp.2016.76
- Cai Y, Dai Y, Wang Y, Yang Q, Guo J, Wei C, Chen W, Huang H, Zhu J, Zhang C, Zheng W, Wen Z, Liu H, Zhang M, Xing S, Jin Q, Feng CG, Chen X (2020). Single-cell transcriptomics of blood reveals a natural killer cell subset depletion in tuberculosis. EBioMedicine 53: 102686. 10.1016/j.ebiom.2020.102686
- Blecher-Gonen R, Bost P, Hilligan KL, David E, Salame TM, Roussel E, Connor LM, Mayer JU, Bahar Halpern K, Tóth B, Itzkovitz S, Schwikowski B, Ronchese F, Amit I (2019). Single-Cell Analysis of Diverse Pathogen Responses Defines a Molecular Roadmap for Generating Antigen-Specific Immunity. Cell Syst 8(2): 109-121.e106. 10.1016/j.cels.2019.01.001
- Li H, Liu SM, Yu XH, Tang SL, Tang CK (2020). Coronavirus disease 2019 (COVID-19): current status and future perspectives. Int J Antimicrob Agents 55(5): 105951. 10.1016/j.ijantimicag.2020.105951
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu N-H, Nitsche A, Müller MA, Drosten C, Pöhlmann S (2020). SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181(2): 271-280.e278. https://doi.org/10.1016/j.cell.2020.02.052
- Walls AC, Park Y-J, Tortorici MA, Wall A, McGuire AT, Veesler D (2020). Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 181(2): 281-292.e286. 10.1016/j.cell.2020.02.058
- Zou X, Chen K, Zou J, Han P, Hao J, Han Z (2020). Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front Med 14(2): 185-192. 10.1007/s11684-020-0754-0
- Xu H, Zhong L, Deng J, Peng J, Dan H, Zeng X, Li T, Chen Q (2020). High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int J Oral Sci 12(1): 8. 10.1038/s41368-020-0074-x
- Sungnak W, Huang N, Bécavin C, Berg M, Queen R, Litvinukova M, Talavera-López C, Maatz H, Reichart D, Sampaziotis F, Worlock KB, Yoshida M, Barnes JL (2020). SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med 26(5): 681-687. 10.1038/s41591-020-0868-6
- Lukassen S, Chua RL, Trefzer T, Kahn NC, Schneider MA, Muley T, Winter H, Meister M, Veith C, Boots AW, Hennig BP, Kreuter M, Conrad C, Eils R (2020). SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J 39(10): e105114. 10.15252/embj.20105114
- Smith JC, Sausville EL, Girish V, Yuan ML, Vasudevan A, John KM, Sheltzer JM (2020). Cigarette Smoke Exposure and Inflammatory Signaling Increase the Expression of the SARS-CoV-2 Receptor ACE2 in the Respiratory Tract. Dev Cell 53(5): 514-529.e513. 10.1016/j.devcel.2020.05.012
- Zheng Y, Liu X, Le W, Xie L, Li H, Wen W, Wang S, Ma S, Huang Z, Ye J, Shi W, Ye Y, Liu Z, Song M, Zhang W, Han J-DJ, Belmonte JCI, Xiao C, Qu J, Wang H, Liu G-H, Su W (2020). A human circulating immune cell landscape in aging and COVID-19. Protein Cell 11(10):740-770. 10.1007/s13238-020-00762-2
- Schuler BA, Habermann AC, Plosa EJ, Taylor CJ, Jetter C, Negretti NM, Kapp ME, Benjamin JT, Gulleman P, Nichols DS, Braunstein LZ, Hackett A, Koval M, Guttentag SH, Blackwell TS, Webber SA, Banovich NE, Kropski JA, Sucre JMS (2020). Age-determined expression of priming protease TMPRSS2 and localization of SARS-CoV-2 in lung epithelium. J Clin Invest 131(1):e140766. 10.1172/jci140766
- Ziegler CGK, Allon SJ, Nyquist SK, Mbano IM, Miao VN, Tzouanas CN, Cao Y, Yousif AS, Bals J, Hauser BM, Feldman J, Muus C, Wadsworth MH, 2nd, Kazer SW, Hughes TK, Doran B, Gatter GJ, Vukovic M, Taliaferro F, Mead BE, Guo Z, Wang JP, Gras D, Plaisant M, Ansari M, Angelidis I, Adler H, Sucre JMS, Taylor CJ, Lin B, et al. (2020). SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 181(5): 1016-1035.e1019. 10.1016/j.cell.2020.04.035
- García LF (2020). Immune Response, Inflammation, and the Clinical Spectrum of COVID-19. Front Immunol 11: 1441. 10.3389/fimmu.2020.01441
- Bost P, Giladi A, Liu Y, Bendjelal Y, Xu G, David E, Blecher-Gonen R, Cohen M, Medaglia C, Li H, Deczkowska A, Zhang S, Schwikowski B, Zhang Z, Amit I (2020). Host-Viral Infection Maps Reveal Signatures of Severe COVID-19 Patients. Cell 181(7): 1475-1488.e1412. 10.1016/j.cell.2020.05.006
- Lee JS, Park S, Jeong HW, Ahn JY, Choi SJ, Lee H, Choi B, Nam SK, Sa M, Kwon JS, Jeong SJ, Lee HK, Park SH, Park SH, Choi JY, Kim SH, Jung I, Shin EC (2020). Immunophenotyping of COVID-19 and influenza highlights the role of type I interferons in development of severe COVID-19. Sci Immunol 5(49). 10.1126/sciimmunol.abd1554
- Schulte-Schrepping J, Reusch N, Paclik D, Baβler K, Schlickeiser S, Zhang B, Krämer B, Krammer T, Brumhard S, Bonaguro L, De Domenico E, Wendisch D, Grasshoff M, Kapellos TS, Beckstette M, Pecht T, Saglam A, Dietrich O, Mei HE, Schulz AR, Conrad C, Kunkel D, Vafadarnejad E, Xu CJ, Horne A, Herbert M, Drews A, Thibeault C, Pfeiffer M, Hippenstiel S, et al. (2020). Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 182(6): 1419-1440.e1423. 10.1016/j.cell.2020.08.001
- Wilk AJ, Rustagi A, Zhao NQ, Roque J, Martínez-Colón GJ, McKechnie JL, Ivison GT, Ranganath T, Vergara R, Hollis T, Simpson LJ, Grant P, Subramanian A, Rogers AJ, Blish CA (2020). A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat Med 26(7): 1070-1076. 10.1038/s41591-020-0944-y
- Liao M, Liu Y, Yuan J, Wen Y, Xu G, Zhao J, Cheng L, Li J, Wang X, Wang F, Liu L, Amit I, Zhang S, Zhang Z (2020). Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat Med 26(6): 842-844. 10.1038/s41591-020-0901-9
- Meckiff BJ, Ramírez-Suástegui C, Fajardo V, Chee SJ, Kusnadi A, Simon H, Eschweiler S, Grifoni A, Pelosi E, Weiskopf D, Sette A, Ay F, Seumois G, Ottensmeier CH, Vijayanand P (2020). Imbalance of Regulatory and Cytotoxic SARS-CoV-2-Reactive CD4(+) T Cells in COVID-19. Cell 183(5):1340-1353.e16. 10.1016/j.cell.2020.10.001
- Zhu L, Yang P, Zhao Y, Zhuang Z, Wang Z, Song R, Zhang J, Liu C, Gao Q, Xu Q, Wei X, Sun H-X, Ye B, Wu Y, Zhang N, Lei G, Yu L, Yan J, Diao G, Meng F, Bai C, Mao P, Yu Y, Wang M, Yuan Y, Deng Q, Li Z, Huang Y, Hu G, Liu Y, et al. (2020). Single-Cell Sequencing of Peripheral Mononuclear Cells Reveals Distinct Immune Response Landscapes of COVID-19 and Influenza Patients. Immunity 53(3):685-696.e3. 10.1016/j.immuni.2020.07.009
- Xu X, Han M, Li T, Sun W, Wang D, Fu B, Zhou Y, Zheng X, Yang Y, Li X, Zhang X, Pan A, Wei H (2020). Effective treatment of severe COVID-19 patients with tocilizumab. Proc Natl Acad Sci U S A 117(20): 10970. 10.1073/pnas.2005615117
- Chua RL, Lukassen S, Trump S, Hennig BP, Wendisch D, Pott F, Debnath O, Thürmann L, Kurth F, Völker MT, Kazmierski J, Timmermann B, Twardziok S, Schneider S, Machleidt F, Müller-Redetzky H, Maier M, Krannich A, Schmidt S, Balzer F, Liebig J, Loske J, Suttorp N, Eils J, Ishaque N, Liebert UG, von Kalle C, Hocke A, Witzenrath M, Goffinet C, et al. (2020). COVID-19 severity correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat Biotechnol 38(8):970-979 10.1038/s41587-020-0602-4
- Daniloski Z, Jordan TX, Wessels HH, Hoagland DA, Kasela S, Legut M, Maniatis S, Mimitou EP, Lu L, Geller E, Danziger O, Rosenberg BR, Phatnani H, Smibert P, Lappalainen T, tenOever BR, Sanjana NE (2020). Identification of Required Host Factors for SARS-CoV-2 Infection in Human Cells. Cell 184(1):92-105.e16. 10.1016/j.cell.2020.10.030
- Cao Y, Su B, Guo X, Sun W, Deng Y, Bao L, Zhu Q, Zhang X, Zheng Y, Geng C, Chai X, He R, Li X, Lv Q, Zhu H, Deng W, Xu Y, Wang Y, Qiao L, Tan Y, Song L, Wang G, Du X, Gao N, Liu J, Xiao J, Su XD, Du Z, Feng Y, Qin C, et al. (2020). Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients’ B Cells. Cell 182(1): 73-84.e16. 10.1016/j.cell.2020.05.025
- Li F, Luo M, Zhou W, Li J, Jin X, Xu Z, Juan L, Zhang Z, Li Y, Liu R, Li Y, Xu C, Ma K, Cao H, Wang J, Wang P, Bu Z, Jiang Q (2020). Single cell RNA and immune repertoire profiling of COVID-19 patients reveal novel neutralizing antibody. Protein Cell 1-5. 10.1007/s13238-020-00807-6
- Hansen J, Baum A, Pascal KE, Russo V, Giordano S, Wloga E, Fulton BO, Yan Y, Koon K, Patel K, Chung KM, Hermann A, Ullman E, Cruz J, Rafique A, Huang T, Fairhurst J, Libertiny C, Malbec M, Lee WY, Welsh R, Farr G, Pennington S, Deshpande D, Cheng J, Watty A, Bouffard P, Babb R, Levenkova N, Chen C, et al. (2020). Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 369(6506): 1010-1014. 10.1126/science.abd0827
- Kazer SW, Aicher TP, Muema DM, Carroll SL, Ordovas-Montanes J, Miao VN, Tu AA, Ziegler CGK, Nyquist SK, Wong EB, Ismail N, Dong M, Moodley A, Berger B, Love JC, Dong KL, Leslie A, Ndhlovu ZM, Ndung’u T, Walker BD, Shalek AK (2020). Integrated single-cell analysis of multicellular immune dynamics during hyperacute HIV-1 infection. Nat Med 26(4): 511-518. 10.1038/s41591-020-0799-2
- Zhang J, Liu J, Yuan Y, Huang F, Ma R, Luo B, Xi Z, Pan T, Liu B, Zhang Y, Zhang X, Luo Y, Wang J, Zhao M, Lu G, Deng K, Zhang H (2020). Two waves of pro-inflammatory factors are released during the influenza A virus (IAV)-driven pulmonary immunopathogenesis. PLoS Pathog 16(2): e1008334. 10.1371/journal.ppat.1008334
- Theel ES, Pritt BS (2016). Parasites. Microbiol Spectr 4(4). 10.1128/microbiolspec.DMIH2-0013-2015
- Phillips MA, Burrows JN, Manyando C, van Huijsduijnen RH, Van Voorhis WC, Wells TNC (2017). Malaria. Nat Rev Dis Primers 3: 17050. 10.1038/nrdp.2017.50
- Josling GA, Llinás M (2015). Sexual development in Plasmodium parasites: knowing when it’s time to commit. Nat Rev Microbiol 13(9): 573-587. 10.1038/nrmicro3519
- Ashley EA, Pyae Phyo A, Woodrow CJ (2018). Malaria. Lancet 391(10130): 1608-1621. 10.1016/s0140-6736(18)30324-6
- Kafsack BF, Rovira-Graells N, Clark TG, Bancells C, Crowley VM, Campino SG, Williams AE, Drought LG, Kwiatkowski DP, Baker DA, Cortés A, Llinás M (2014). A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 507(7491): 248-252. 10.1038/nature12920
- Poran A, Nötzel C, Aly O, Mencia-Trinchant N, Harris CT, Guzman ML, Hassane DC, Elemento O, Kafsack BFC (2017). Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites. Nature 551(7678): 95-99. 10.1038/nature24280
- Reid AJ, Talman AM, Bennett HM, Gomes AR, Sanders MJ, Illingworth CJR, Billker O, Berriman M, Lawniczak MK (2018). Single-cell RNA-seq reveals hidden transcriptional variation in malaria parasites. Elife 7:e33105. 10.7554/eLife.33105
- Walzer KA, Fradin H, Emerson LY, Corcoran DL, Chi J-T (2019). Latent transcriptional variations of individual Plasmodium falciparum uncovered by single-cell RNA-seq and fluorescence imaging. PLoS Genet 15(12): e1008506-e1008506. 10.1371/journal.pgen.1008506
- Bancells C, Llorà-Batlle O, Poran A, Nötzel C, Rovira-Graells N, Elemento O, Kafsack BFC, Cortés A (2019). Revisiting the initial steps of sexual development in the malaria parasite Plasmodium falciparum. Nat Microbiol 4(1): 144-154. 10.1038/s41564-018-0291-7
- Ngara M, Palmkvist M, Sagasser S, Hjelmqvist D, Björklund Å K, Wahlgren M, Ankarklev J, Sandberg R (2018). Exploring parasite heterogeneity using single-cell RNA-seq reveals a gene signature among sexual stage Plasmodium falciparum parasites. Exp Cell Res 371(1): 130-138. 10.1016/j.yexcr.2018.08.003
- Howick VM, Russell AJC, Andrews T, Heaton H, Reid AJ, Natarajan K, Butungi H, Metcalf T, Verzier LH, Rayner JC, Berriman M, Herren JK, Billker O, Hemberg M, Talman AM, Lawniczak MKN (2019). The Malaria Cell Atlas: Single parasite transcriptomes across the complete Plasmodium life cycle. Science 365(6455): eaaw2619. 10.1126/science.aaw2619
- Kumar R, Loughland JR, Ng SS, Boyle MJ, Engwerda CR (2020). The regulation of CD4(+) T cells during malaria. Immunol Rev 293(1): 70-87. 10.1111/imr.12804
- Smith NL, Nahrendorf W, Sutherland C, Mooney JP, Thompson J, Spence PJ, Cowan GJM (2020). A Conserved TCRβ Signature Dominates a Highly Polyclonal T-Cell Expansion During the Acute Phase of a Murine Malaria Infection. Front Immunol 11: 587756. 10.3389/fimmu.2020.587756
- Perez-Mazliah D, Langhorne J (2015). CD4 T-Cell Subsets in Malaria: TH1/TH2 Revisited. Front Immunol 5: 671. 10.3389/fimmu.2014.00671
- Obeng-Adjei N, Portugal S, Tran Tuan M, Yazew Takele B, Skinner J, Li S, Jain A, Felgner Philip L, Doumbo Ogobara K, Kayentao K, Ongoiba A, Traore B, Crompton Peter D (2015). Circulating Th1-Cell-type Tfh Cells that Exhibit Impaired B Cell Help Are Preferentially Activated during Acute Malaria in Children. Cell Rep 13(2): 425-439. 10.1016/j.celrep.2015.09.004
- Lönnberg T, Svensson V, James KR, Fernandez-Ruiz D, Sebina I, Montandon R, Soon MS, Fogg LG, Nair AS, Liligeto U, Stubbington MJ, Ly LH, Bagger FO, Zwiessele M, Lawrence ND, Souza-Fonseca-Guimaraes F, Bunn PT, Engwerda CR, Heath WR, Billker O, Stegle O, Haque A, Teichmann SA (2017). Single-cell RNA-seq and computational analysis using temporal mixture modelling resolves Th1/Tfh fate bifurcation in malaria. Sci Immunol 2(9): eaal2192. 10.1126/sciimmunol.aal2192
- Fontana MF, de Melo GL, Anidi C, Hamburger R, Kim CY, Lee SY, Pham J, Kim CC (2016). Macrophage Colony Stimulating Factor Derived from CD4+ T Cells Contributes to Control of a Blood-Borne Infection. PLoS Pathog 12(12): e1006046. 10.1371/journal.ppat.1006046
- Cockburn IA, Seder RA (2018). Malaria prevention: from immunological concepts to effective vaccines and protective antibodies. Nat Immunol 19(11): 1199-1211. 10.1038/s41590-018-0228-6
- Bongomin F, Gago S, Oladele RO, Denning DW (2017). Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J Fungi 3(4): 57. 10.3390/jof3040057
- Suleyman G, Alangaden GJ (2016). Nosocomial Fungal Infections: Epidemiology, Infection Control, and Prevention. Infect Dis Clin North Am 30(4): 1023-1052. 10.1016/j.idc.2016.07.008
- Vazquez JA, Miceli MH, Alangaden G (2013). Invasive fungal infections in transplant recipients. Ther Adv Infect Dis1(3): 85-105. 10.1177/2049936113491936
- Kotthoff-Burrell E (2019). Candidemia (Blood Infection) and Other Candida Infections. Am J Respir Crit Care Med 200(5): P9-p10. 10.1164/rccm.2005P9
- de Vries DH, Matzaraki V, Bakker OB, Brugge H, Westra HJ, Netea MG, Franke L, Kumar V, van der Wijst MGP (2020). Integrating GWAS with bulk and single-cell RNA-sequencing reveals a role for LY86 in the anti-Candida host response. PLoS Pathog 16(4): e1008408. 10.1371/journal.ppat.1008408
- de Bekker C, Bruning O, Jonker MJ, Breit TM, Wösten HA (2011). Single cell transcriptomics of neighboring hyphae of Aspergillus niger. Genome Biol 12(8): R71. 10.1186/gb-2011-12-8-r71
- Ku C, Sebé-Pedrós A (2019). Using single-cell transcriptomics to understand functional states and interactions in microbial eukaryotes. Philos Trans R Soc Lond B Biol Sci 374(1786): 20190098. 10.1098/rstb.2019.0098
- Penaranda C, Hung DT (2019). Single-Cell RNA Sequencing to Understand Host-Pathogen Interactions. ACS Infect Dis 5(3): 336-344. 10.1021/acsinfecdis.8b00369
- Lähnemann D, Köster J, Szczurek E, McCarthy DJ, Hicks SC, Robinson MD, Vallejos CA, Campbell KR, Beerenwinkel N, Mahfouz A, Pinello L, Skums P, Stamatakis A, Attolini CS, Aparicio S, Baaijens J, Balvert M, Barbanson B, Cappuccio A, Corleone G, Dutilh BE, Florescu M, Guryev V, Holmer R, Jahn K, Lobo TJ, Keizer EM, Khatri I, Kielbasa SM, Korbel JO, et al. (2020). Eleven grand challenges in single-cell data science. Genome Biol 21(1): 31. 10.1186/s13059-020-1926-6
- Chen M, Zhou X (2018). VIPER: variability-preserving imputation for accurate gene expression recovery in single-cell RNA sequencing studies. Genome Biol 19(1): 196. 10.1186/s13059-018-1575-1
- Chi W, Deng M (2020). Sparsity-Penalized Stacked Denoising Autoencoders for Imputing Single-Cell RNA-Seq Data. Genes 11(5): 532. 10.3390/genes11050532
- Westermann AJ, Forstner KU, Amman F, Barquist L, Chao Y, Schulte LN, Muller L, Reinhardt R, Stadler PF, Vogel J (2016). Dual RNA-seq unveils noncoding RNA functions in host-pathogen interactions. Nature 529(7587): 496-501. 10.1038/nature16547
- Turchinovich A, Surowy H, Serva A, Zapatka M, Lichter P, Burwinkel B (2014). Capture and Amplification by Tailing and Switching (CATS). An ultrasensitive ligation-independent method for generation of DNA libraries for deep sequencing from picogram amounts of DNA and RNA. RNA Biol 11(7): 817-828. 10.4161/rna.29304
- Shishkin AA, Giannoukos G, Kucukural A, Ciulla D, Busby M, Surka C, Chen J, Bhattacharyya RP, Rudy RF, Patel MM, Novod N, Hung DT, Gnirke A, Garber M, Guttman M, Livny J (2015). Simultaneous generation of many RNA-seq libraries in a single reaction. Nat Methods 12(4): 323-325. 10.1038/nmeth.3313
- Lafzi A, Moutinho C, Picelli S, Heyn H (2018). Tutorial: guidelines for the experimental design of single-cell RNA sequencing studies. Nat Protoc 13(12): 2742-2757. 10.1038/s41596-018-0073-y
- Raj A, van den Bogaard P, Rifkin SA, van Oudenaarden A, Tyagi S (2008). Imaging individual mRNA molecules using multiple singly labeled probes. Nat Methods 5(10): 877-879. 10.1038/nmeth.1253
- Chen KH, Boettiger AN, Moffitt JR, Wang S, Zhuang X (2015). Spatially resolved, highly multiplexed RNA profiling in single cells. Science 348(6233): aaa6090. 10.1126/science.aaa6090
- Eng C-HL, Lawson M, Zhu Q, Dries R, Koulena N, Takei Y, Yun J, Cronin C, Karp C, Yuan G-C, Cai L (2019). Transcriptome-scale super-resolved imaging in tissues by RNA seqFISH+. Nature 568(7751): 235-239. 10.1038/s41586-019-1049-y
- Moncada R, Barkley D, Wagner F, Chiodin M, Devlin JC, Baron M, Hajdu CH, Simeone DM, Yanai I (2020). Integrating microarray-based spatial transcriptomics and single-cell RNA-seq reveals tissue architecture in pancreatic ductal adenocarcinomas. Nat Biotechnol 38(3): 333-342. 10.1038/s41587-019-0392-8
- Edsgärd D, Johnsson P, Sandberg R (2018). Identification of spatial expression trends in single-cell gene expression data. Nat Methods 15(5): 339-342. 10.1038/nmeth.4634
- Trapnell C, Cacchiarelli D, Grimsby J, Pokharel P, Li S, Morse M, Lennon NJ, Livak KJ, Mikkelsen TS, Rinn JL (2014). The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat Biotechnol 32(4): 381-386. 10.1038/nbt.2859
- Abbas A, Vu Manh T-P, Valente M, Collinet N, Attaf N, Dong C, Naciri K, Chelbi R, Brelurut G, Cervera-Marzal I, Rauwel B, Davignon J-L, Bessou G, Thomas-Chollier M, Thieffry D, Villani A-C, Milpied P, Dalod M, Tomasello E (2020). The activation trajectory of plasmacytoid dendritic cells in vivo during a viral infection. Nat Immunol 21(9): 983-997. 10.1038/s41590-020-0731-4
- Khatun A, Kasmani MY, Zander R, Schauder DM, Snook JP, Shen J, Wu X, Burns R, Chen YG, Lin CW, Williams MA, Cui W (2021). Single-cell lineage mapping of a diverse virus-specific naive CD4 T cell repertoire. J Exp Med 218(3): e20200650. 10.1084/jem.20200650
- Saelens W, Cannoodt R, Todorov H, Saeys Y (2019). A comparison of single-cell trajectory inference methods. Nat Biotechnol 37(5): 547-554. 10.1038/s41587-019-0071-9
- Gawel DR, Serra-Musach J, Lilja S, Aagesen J, Arenas A, Asking B, Bengnér M, Björkander J, Biggs S, Ernerudh J, Hjortswang H, Karlsson JE, Köpsen M, Lee EJ, Lentini A, Li X, Magnusson M, Martínez-Enguita D, Matussek A, Nestor CE, Schäfer S, Seifert O, Sonmez C, Stjernman H, Tjärnberg A, Wu S, Åkesson K, Shalek AK, Stenmarker M, Zhang H, et al. (2019). A validated single-cell-based strategy to identify diagnostic and therapeutic targets in complex diseases. Genome Med 11(1): 47. 10.1186/s13073-019-0657-3
- Bossel Ben-Moshe N, Hen-Avivi S, Levitin N, Yehezkel D, Oosting M, Joosten LAB, Netea MG, Avraham R (2019). Predicting bacterial infection outcomes using single cell RNA-sequencing analysis of human immune cells. Nat Commun 10(1): 3266. 10.1038/s41467-019-11257-y
- Alakwaa FM (2020). Repurposing Didanosine as a Potential Treatment for COVID-19 Using Single-Cell RNA Sequencing Data. mSystems 5(2): e00297-00220. 10.1128/mSystems.00297-20
- Rusk N (2019). Multi-omics single-cell analysis. Nat Methods 16(8): 679. 10.1038/s41592-019-0519-3
- Rooijers K, Markodimitraki CM, Rang FJ, de Vries SS, Chialastri A, de Luca KL, Mooijman D, Dey SS, Kind J (2019). Simultaneous quantification of protein-DNA contacts and transcriptomes in single cells. Nat Biotechnol 37(7): 766-772. 10.1038/s41587-019-0150-y
- Ma S, Zhang B, LaFave LM, Earl AS, Chiang Z, Hu Y, Ding J, Brack A, Kartha VK, Tay T, Law T, Lareau C, Hsu YC, Regev A, Buenrostro JD (2020). Chromatin Potential Identified by Shared Single-Cell Profiling of RNA and Chromatin. Cell 183(4): 1103-1116.e1120. 10.1016/j.cell.2020.09.056
- Bumann D (2015). Heterogeneous host-pathogen encounters: act locally, think globally. Cell Host Microbe 17(1): 13-19. 10.1016/j.chom.2014.12.006
- Zheng Y, Liu X, Le W, Xie L, Li H, Wen W, Wang S, Ma S, Huang Z, Ye J, Shi W, Ye Y, Liu Z, Song M, Zhang W, Han JJ, Belmonte JCI, Xiao C, Qu J, Wang H, Liu GH, Su W (2020). A human circulating immune cell landscape in aging and COVID-19. Protein Cell 11(10): 740-770. 10.1007/s13238-020-00762-2
- Steuerman Y, Cohen M, Peshes-Yaloz N, Valadarsky L, Cohn O, David E, Frishberg A, Mayo L, Bacharach E, Amit I, Gat-Viks I (2018). Dissection of Influenza Infection In Vivo by Single-Cell RNA Sequencing. Cell Syst 6(6): 679-691.e674. 10.1016/j.cels.2018.05.008
- Ramos I, Smith G, Ruf-Zamojski F, Martínez-Romero C, Fribourg M, Carbajal EA, Hartmann BM, Nair VD, Marjanovic N, Monteagudo PL, DeJesus VA, Mutetwa T, Zamojski M, Tan GS, Jayaprakash C, Zaslavsky E, Albrecht RA, Sealfon SC, García-Sastre A, Fernandez-Sesma A (2019). Innate Immune Response to Influenza Virus at Single-Cell Resolution in Human Epithelial Cells Revealed Paracrine Induction of Interferon Lambda 1. J Virol 93(20): e00559-00519. 10.1128/JVI.00559-19
- Wang C, Forst CV, Chou TW, Geber A, Wang M, Hamou W, Smith M, Sebra R, Zhang B, Zhou B, Ghedin E (2020). Cell-to-Cell Variation in Defective Virus Expression and Effects on Host Responses during Influenza Virus Infection. mBio 11(1): e02880-19. 10.1128/mBio.02880-19
- Rato S, Rausell A, Muñoz M, Telenti A, Ciuffi A (2017). Single-cell analysis identifies cellular markers of the HIV permissive cell. PLoS Pathog 13(10): e1006678. 10.1371/journal.ppat.1006678
- Bradley T, Ferrari G, Haynes BF, Margolis DM, Browne EP (2018). Single-Cell Analysis of Quiescent HIV Infection Reveals Host Transcriptional Profiles that Regulate Proviral Latency. Cell Rep 25(1): 107-117.e103. 10.1016/j.celrep.2018.09.020
- Golumbeanu M, Cristinelli S, Rato S, Munoz M, Cavassini M, Beerenwinkel N, Ciuffi A (2018). Single-Cell RNA-Seq Reveals Transcriptional Heterogeneity in Latent and Reactivated HIV-Infected Cells. Cell Rep 23(4): 942-950. 10.1016/j.celrep.2018.03.102
- Onorati M, Li Z, Liu F, Sousa André MM, Nakagawa N, Li M, Dell’Anno Maria T, Gulden Forrest O, Pochareddy S, Tebbenkamp Andrew TN, Han W, Pletikos M, Gao T, Zhu Y, Bichsel C, Varela L, Szigeti-Buck K, Lisgo S, Zhang Y, Testen A, Gao X-B, Mlakar J, Popovic M, Flamand M, Strittmatter Stephen M, Kaczmarek Leonard K, Anton ES, Horvath Tamas L, Lindenbach Brett D, Sestan N (2016). Zika Virus Disrupts Phospho-TBK1 Localization and Mitosis in Human Neuroepithelial Stem Cells and Radial Glia. Cell Rep 16(10): 2576-2592. 10.1016/j.celrep.2016.08.038
- Nowakowski Tomasz J, Pollen Alex A, Di Lullo E, Sandoval-Espinosa C, Bershteyn M, Kriegstein Arnold R (2016). Expression Analysis Highlights AXL as a Candidate Zika Virus Entry Receptor in Neural Stem Cells. Cell Stem Cell 18(5): 591-596. 10.1016/j.stem.2016.03.012
- Gorman MJ, Caine EA, Zaitsev K, Begley MC, Weger-Lucarelli J, Uccellini MB, Tripathi S, Morrison J, Yount BL, Dinnon KH, 3rd, Rückert C, Young MC, Zhu Z, Robertson SJ, McNally KL, Ye J, Cao B, Mysorekar IU, Ebel GD, Baric RS, Best SM, Artyomov MN, Garcia-Sastre A, Diamond MS (2018). An Immunocompetent Mouse Model of Zika Virus Infection. Cell Host Microbe 23(5): 672-685.e676. 10.1016/j.chom.2018.04.003
- Zanini F, Robinson ML, Croote D, Sahoo MK, Sanz AM, Ortiz-Lasso E, Albornoz LL, Rosso F, Montoya JG, Goo L, Pinsky BA, Quake SR, Einav S (2018). Virus-inclusive single-cell RNA sequencing reveals the molecular signature of progression to severe dengue. Proc Natl Acad Sci U S A 115(52): E12363-e12369. 10.1073/pnas.1813819115
- Kotliar D, Lin AE, Logue J, Hughes TK, Khoury NM, Raju SS, Wadsworth MH, Chen H, Kurtz JR, Dighero-Kemp B, Bjornson ZB, Mukherjee N, Sellers BA, Tran N, Bauer MR, Adams GC, Adams R, Rinn JL, Melé M, Schaffner SF, Nolan GP, Barnes KG, Hensley LE, McIlwain DR, Shalek AK, Sabeti PC, Bennett RS (2020). Single-Cell Profiling of Ebola Virus Disease In Vivo Reveals Viral and Host Dynamics. Cell 183(5): 1383-1401.e1319. 10.1016/j.cell.2020.10.002
- Walzer KA, Fradin H, Emerson LY, Corcoran DL, Chi JT (2019). Latent transcriptional variations of individual Plasmodium falciparum uncovered by single-cell RNA-seq and fluorescence imaging. PLoS Genet 15(12): e1008506. 10.1371/journal.pgen.1008506
- Vigneron A, O’Neill MB, Weiss BL, Savage AF, Campbell OC, Kamhawi S, Valenzuela JG, Aksoy S (2020). Single-cell RNA sequencing of Trypanosoma brucei from tsetse salivary glands unveils metacyclogenesis and identifies potential transmission blocking antigens. Proc Natl Acad Sci U S A 117(5): 2613-2621. 10.1073/pnas.1914423117
- Patir A, Gossner A, Ramachandran P, Alves J, Freeman TC, Henderson NC, Watson M, Hassan MA (2020). Single-cell RNA-seq reveals CD16(-) monocytes as key regulators of human monocyte transcriptional response to Toxoplasma. Sci Rep 10(1): 21047. 10.1038/s41598-020-78250-0
–
ACKNOWLEDGMENTS
This work was supported by grants from the National Natural Science Foundation of China (No. 81830068, No. 81772140, No. 31700120, and No. 81501733), Key Research and Development Project of China (No. 2016YFA0500600), the Program for Professor of Special Appointment (Eastern Scholar) at Shanghai Institutions of Higher Learning.
COPYRIGHT
© 2021

Understanding the pathogenesis of infectious diseases by single-cell RNA sequencing by Huang et al. is licensed under a Creative Commons Attribution 4.0 International License.








