
Editorial:
Microbial Cell, Vol. 9, No. 4, pp. 72 - 79; doi: 10.15698/mic2022.04.773
A hundred spotlights on microbiology: how microorganisms shape our lives

1 Institute of Molecular Biosciences, NAWI Graz, University of Graz, Graz, Austria.
2 Centre de Recherche des Cordeliers, Equipe labellisée par la Ligue contre le cancer, Université de Paris Cité, Sorbonne Université, Inserm U1138, Institut Universitaire de France, Paris, France.
3 Metabolomics and Cell Biology Platforms, Institut Gustave Roussy, Villejuif, France.
4 Institut du Cancer Paris CARPEM, Department of Biology, Hôpital Européen Georges Pompidou, AP-HP, Paris, France.
5 BioHealth Graz, Graz, Austria.
6 BioTechMed Graz, Graz 8010, Austria.
Keywords: infectious diseases, model organism, yeast, biotechnology, microbiome, microbiota, symbiosis, open access.
Received originally: 11/03/2022 Accepted: 22/03/2022
Published: 04/04/2022
Correspondence:
Frank Madeo, Humboldtstrasse 50/EG, 8010 Graz; frank.madeo@uni-graz.at
Didac Carmona-Gutierrez, Humboldtstrasse 50/EG, 8010 Graz; carmonag@uni-graz.at
Conflict of interest statement: No competing financial interests exist.
Please cite this article as: Didac Carmona-Gutierrez, Katharina Kainz, Andreas Zimmermann, Sebastian J. Hofer, Maria A. Bauer, Christoph Ruckenstuhl, Guido Kroemer and Frank Madeo (2022). A hundred spotlights on microbiology: how microorganisms shape our lives. Microbial Cell 9(4): 72-79. doi: 10.15698/mic2022.04.773
Viral, bacterial, fungal and protozoal biology is of cardinal importance for the evolutionary history of life, ecology, biotechnology and infectious diseases. Various microbiological model systems have fundamentally contributed to the understanding of molecular and cellular processes, including the cell cycle, cell death, mitochondrial biogenesis, vesicular fusion and autophagy, among many others. Microbial interactions within the environment have profound effects on many fields of biology, from ecological diversity to the highly complex and multifaceted impact of the microbiome on human health. Also, biotechnological innovation and corresponding industrial operations strongly depend on microbial engineering. With this wide range of impact in mind, the peer-reviewed and open access journal Microbial Cell was founded in 2014 and celebrates its 100th issue this month. Here, we briefly summarize how the vast diversity of microbiological subjects influences our personal and societal lives and shortly review the milestones achieved by Microbial Cell during the last years.
THE MANY IMPLICATIONS AND CHALLENGES OF MICROBIAL RESEARCH
The history of life on Earth is mainly microbial. The emergence of the first microorganisms 3-4 billion years ago [1] was the initial step for the establishment of terrestrial life. Microorganisms critically contributed to our planet’s transformation, with the rise of photosynthetic bacteria allowing for oxygen to build up in the atmosphere [2]. Nowadays, microorganisms continue to affect the planet’s biosphere and are an integral and inextricable part of our lives at different levels. The exploration of the microbial world is not only key to understanding ourselves but can provide answers to many medical, technological and scientific questions we face as humankind. Here, we briefly summarize the main areas, on which microorganisms impact today and will in the future, in particular (i) infectious diseases, (ii) symbiotic interactions, (iii) biotechnological applications and (iv) biological models (Figure 1).
–
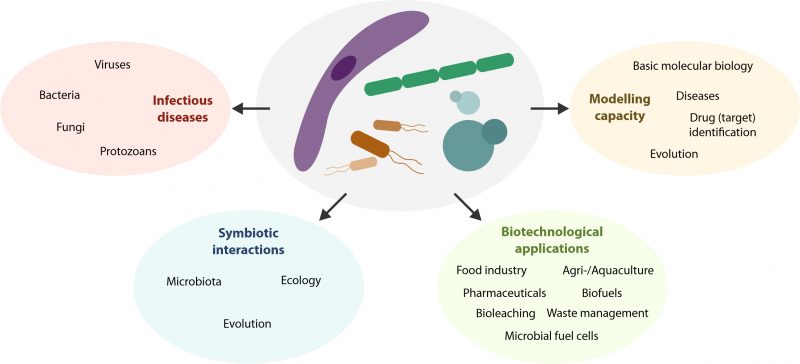 | FIGURE 1: The multilayered impact of microorganisms on our lives. |
INFECTIOUS DISEASES
Infectious diseases continue to threaten our lives and societies, as revealed by the current COVID-19 pandemic. This applies in particular to persisting, emerging and re-emerging infectious diseases. In that respect, it will be important to monitor the human-animal interface, and recognize that many emerging diseases are zoonotic, i.e. they spread between animals and humans and have an animal reservoir [3]. This threat emanates from all microbial phyla. Apart from minatory viral infections [4], bacterial and fungal infections continue to cause millions of deaths worldwide [5][6]. An alarming rise in antibiotic and antifungal resistance exacerbates this menace [5][7], representing one of the most acute challenges in medical microbiology. Finally, protozoan infections remain a public health threat associated with significant morbidity and continue to have a substantial socioeconomic impact [8][9]. The limited advancements in vaccination strategies and the increasing parasite resistance against existing drugs further exacerbate the problem [10]. Also, coinfections of different pathogens belonging to one or different species can occur, resulting in additional challenges [11][12][13][14]. Thus, more work is needed to explore the molecular and pathological interaction between co-infecting microorganisms. This applies to both the direct interplay with each other and the indirect interaction through the host, for instance via the immune system. Altogether, infectious diseases continue to be devastating despite the many medical improvements accomplished in the last decades. Of note, many microbial pathogens also pose a threat to agriculture and livestock farming [15][16], which further aggravates the socioeconomic burden of infectious diseases.
SYMBIOTIC INTERACTIONS
The participation of microorganisms in symbiotic interactions determines a vast range of biological aspects across species. In particular, the human gut microbiota – the collection of bacteria, archaea and eukarya colonizing the gastrointestinal tract [17] – has attracted much attention in the past decade. The gut microbiota has a deep impact on many instances of human biology, ranging from development, physiology and immune homeostasis to health, nutrition and even behavior [18][19][20][21]. In addition, the importance of the human virome is increasingly being recognized. The human virome is composed of bacteriophages that infect bacteria, viruses that infect other cellular microorganisms (archaea, eukarya), viruses that infect human cells and some transitory viruses originating in food [22]. Their interactions with the human host are only beginning to be understood, but clearly hint towards a decisive role in health (e.g. via interplay with the host immune system) and multiple diseases (e.g. diabetes, hypertension and cancer) [22]. From a broader perspective, microbial symbiosis affects and co-defines a vast array of ecological aspects, ranging from plant growth [23] to defensive capacities [24]. For instance, complete ecosystems like hydrothermal vents and coral reefs exemplify the ecological success of microbial-multicellular symbioses. Finally, symbiosis can uniquely drive evolutionary innovation; this is maybe best embodied in the endosymbiotic origins of mitochondria and chloroplasts in eukaryotic cells [25][26], where the symbiont was cellularly and genomically integrated into the host.
BIOTECHNOLOGY
From a historical or even pre-historical perspective, already in ancient times, microorganisms were used as tools to produce, ferment or process a diversity of important food items including vinegar, bread, beer, fish, cheese and wine [27]. In other words, microorganisms were actively used as biotechnological agents long before the scientific basis underlying these processes was even known. Nowadays, microorganisms represent an essential backbone of many biotechnological applications thanks to their rapid growth for quick production, technical versatility for production design and wide applicability to a number of industrial sectors. Indeed, fundamental and applied microbiology are essential components of modern biotechnology with an ever-increasing economic impact. For instance, the food industry heavily relies on microorganisms for applications that range from fermented food items and alcoholic beverages to food grade components and bio-based ingredients in general. Thus, bacteria are used for the production of thickening or gelling agents, flavor compounds and enhancers, acidulants, vitamins and colorants [28]. Moreover, microorganisms have a deep impact on biotechnological approaches in agriculture and aquaculture with developments ahead that may use host-microbe interactions and the host microbiome for sustainable production [29]. Microorganisms also play a significant role in environmental biotechnology, including municipal and industrial water waste management [30] as well as treatment of solid hazardous waste [31]. Other applications involve the treatment of oil spillage [32], radioactive contamination [33], electronic waste processing [34], bioleaching (the extraction of metals from their ores through the action of microorganisms) [35] or even space biomining [36]. Yet another economically relevant use is energy production. Biofuels are produced by engineered microorganisms that utilize renewable carbon sources. Although they have shown great potential in replacing fossil fuels (especially ethanol and biodiesel), there are still some limitations, including applicability in conventional engines and high costs [37]. So-called microbial fuel cells (MFCs), which use bacteria to oxidize organic and inorganic matter in order to generate current, may represent an appealing electrogenic approach in the future [38]. Another example of microbial biotechnology is pharmaceutical production, which includes heterologous expression of human proteins, microbial enzymes or drug compounds for medical and research purposes [39]. Some of the aforementioned examples are established processes while others are still in development, revealing the huge potential and economic impact of microorganisms in technological approaches [40].
MODELLING BASIC PRINCIPLES OF BIOLOGY
The short generation time, facile cultivation and ease of genetic manipulation have established a number of microorganisms as widely used model organisms. Escherichia coli has been instrumental in the discovery and understanding of basic molecular biology processes, including DNA replication, DNA-to-RNA transcription and the genetic code allowing for RNA-to-protein translation. To date, a total of twelve Nobel Prizes were awarded for work that used E. coli as a research organism or tool, and its potential to assist in further advancements remains high [41]. A number of other prokaryotes (including archaea [42]) are actively used as model organisms. For example, Bacillus subtilis is applied to study biofilms, bacterial asymmetry or morphogenesis [43], cyanobacteria like Synechocystis sp. PCC 6803 to model photosynthesis [44], or Caulobacter crescentus for the study of cellular differentiation, motility or mechanosensing [45]. Green algae, including unicellular Chlamydomonas reinhardtii and multicellular Volvox carteri (Volvox), also serve as model organisms. C. reinhardtii bears both animal-like organelles (cilia) and plant-like organelles (chloroplasts), allowing research into the function of flagella and photosynthesis [46]. Volvox is mainly used to investigate developmental mechanisms and the evolutionary origins of multicellularity [47]. Other protists include the giant heterotrichous ciliate Stentor coeruleus as a model for cellular regeneration and wound healing [48] and the ciliate Oxytricha, which is employed in the areas of genome biology, post-zygotic development and epigenetic inheritance [49]. The eukaryotic nature of these cells allows for the study of essential and medically relevant molecular processes. Similarly, yeast cells display all advantages of unicellular model organisms paired with a high degree of conservation that has made yeast a fundamental partner in elucidating many aspects of human physiology and pathology [50]. Work performed in yeast has been awarded five Nobel Prizes in the past two decades. The budding yeast Saccharomyces cerevisiae is used to study a multitude of human diseases (e.g. neurodegeneration, cancer) [51][52], characterize basic physiological processes (e.g. cell death, aging, autophagy, mitochondrial import, vesicle fusion, cell cycle) [53][54][55][56][57][58][59][60][61][62][63][64] and identify novel medical drugs (e.g. antiaging, anticancer, antiparasitic, antifungal) [56][65][66][67][68][69][70], among many other applications [71]. Another example is the fission yeast Schizosaccharomyces pombe, which is used to analyze, for example, cell cycle processes or DNA checkpoints [72][73].
A PUBLICATION PLATFORM FOR MICROBIOLOGICAL RESEARCH
Given the many layers of how microorganisms wield huge influence on our lives, the open-access journal Microbial Cell was founded with the idea to generate an online agora for all types of research in the microbiological field. The current issue (Volume 9, Issue 4) marks a milestone in Microbial Cell‘s history: it represents the 100th issue since the journal was launched in January 2014. This occasion is a timely moment to take stock and reflect on how Microbial Cell has developed and contributed to the research fields of unicellular and multicellular microorganisms over the past years.
–
The mere fact that a journal run by active academic scientists and through an independent publisher (Shared Science Publishers) has established itself in the highly competitive business of peer-reviewed scientific publishing is per se a great achievement. That this has occurred in a radically open-access fashion and in such an important and trending field like microbiology adds even more value to this accomplishment. We take this opportunity to thank all authors who have published their articles in Microbial Cell for their trust put in the journal to run the evaluation and dissemination processes of their work. At the same time, we congratulate the authors for the high quality of their papers: the around 400 articles that have been published since the journal was launched have now cumulatively been cited more than 4000 times (Web of Science, Clarivate Analytics).
–
We acknowledge all members of the Editorial Board for their long-standing commitment and reliability. Indeed, our Editorial Board has the arduous task to evaluate submissions in ten different thematic subareas: aging, cell death, cell physiology and cell signaling, genome stability and structure, infection biology, microbiome, mitochondria, parasitology, stress response, and structural & systems biology. During the manuscript evaluation process, these editors strongly rely on the expertise and rigor of peer reviewers, who invest a great amount of their time in improving the submitted work. We are very grateful to all our referees throughout the world for their invaluable input. The combined effort of all partners of the ecosystem, authors, editors and reviewers, has consistently improved the journal throughout these years to place it at the apex of microbiology.
A BRIEF HISTORY OF MICROBIAL CELL
Microbial Cell emerged as an academic effort from a group of active scientists to apprehend the thematic heterogeneity of microbial research. Accordingly, the inaugural Editorial from January 2014 outlined that Microbial Cell would have the mission to facilitate “the characterization of unicellular organisms (or multicellular microorganisms) in their response to internal and external stimuli and/or in the context of human health and disease” [74]. This definition mirrored well the initial and still persisting idea of a publication platform that acknowledges microbiological interdisciplinarity.
–
Following these objectives, the first 100 issues of Microbial Cell have accompanied and contributed to several developments in microbiological research during the last eight years. For instance, in a much regarded Microbial Cell paper of 2015, Alexander Varshavsky and colleagues described formylated N-terminal methionine as a novel bacterial degradation signal used in a new branch of the bacterial N-end rule pathway [75]. Varshavsky’s lab was instrumental in the discovery of the ubiquitin system of intracellular protein degradation and was the first to describe the connection between the N-terminal residue of a protein and its half-life [76][77]. As another example, in 2016 Microbial Cell published a highly cited review, in which Daniel J. Klionsky and colleagues provided a brief but comprehensive summary on the roles, regulatory instances and molecular mechanisms of autophagy [78]. 2016 was also the year, in which Yoshinori Ohsumi was awarded the Nobel Prize in Physiology or Medicine for his discovery of the mechanisms orchestrating this intracellular degradation pathway, which were uncovered in yeast cells [79][80][81]. In 2018 Microbial Cell published a review by Francisco J.M. Mójica and colleagues that provided an overview of the CRISPR-Cas mechanism as a prokaryotic immune system and discussed a number of evolutionary implications [82]. Mójica’s achievements, particularly his groundbreaking work characterizing CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) loci [83][84], laid the ground for the development of CRISPR-Cas as the most important tool for genomic editing that exists to date. In 2020, Emmanuelle Charpentier and Jennifer A. Doudna received the Nobel Prize in Chemistry for the development of this method [85]. One final example for how Microbial Cell has contributed to research deals with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which causes the highly infectious disease COVID-19. The multilayered consequences of COVID-19 at the individual, social, political and economic levels are unprecedented in our globalized world. Microbial Cell has been publishing diverse papers on SARS-CoV-2, ranging from mechanistic viewpoints to methodological approaches for viral detection and molecular modelling.
–
The last 100 issues have seen Microbial Cell grow as a journal and establish itself in the microbiology publishing sphere. The journal’s CiteScore (Scopus) is currently 7.1 (provisional score for 2021), which ranks the journal at top positions in all relevant microbiology-related categories. Microbial Cell has established roots in various research communities as highlighted by reviews, research papers, commentaries, as well as by a number of guideline papers that establish unified criteria in a number of fields, for instance in DNA recombination and repair, yeast cell death and antifungal and antibiofilm agents. Furthermore, Microbial Cell has initiated the publication of several Special Issues on a diverse range of topics, following a concept, in which incoming papers are published in regular issues and then collected in the Special Issue repositorium. That way, Special Issues can be updated continuously and refreshed by new articles that capture new evolving knowledge. Until now, Microbial Cell has launched Special Issues on the following topics [86]: sexually transmitted infections (started in 2016), the human microbiome in health and disease (started in 2019), hygiene in healthcare (started in 2019), microbiology in cultural heritage (started in 2021) and yeast cell death (started in 2022).
–
In the course of the last eight years, Microbial Cell has strengthened its position in the microbiology-publishing landscape by attaining a number of milestones. In 2014, Microbial Cell partnered with the World Health Organization’s HINARI program to support free access to biomedical research literature. That same year, Microbial Cell also became a member of Crossref, an official digital object identifier (DOI) registration agency, providing each article with a persistent interoperable identifier that also enables to precisely link citations across publishers of online academic journals. Also in 2014, Microbial Cell was indexed in Sherpa/RoMEO, an aggregator of open access policies of academic journals. In addition, Microbial Cell secured a partnership with the Austrian National Library to ensure digital long-term archiving and perpetual access to its complete content. After successful applications to the Chemical Abstracts Service (CAS) and the Directory of Open Access Journals (DOAJ), Microbial Cell was accepted in these two renowned repositories in 2015. One year later, Microbial Cell was awarded the very selective DOAJ Seal (allocated to only 10% of DOAJ-indexed journals) for best practice in open access publishing. Also in 2016, Microbial Cell entered the International Committee of Medical Journal Editors (ICMJE) list to acknowledge that it follows the ICMJE’s Recommendations for the Conduct, Reporting, Editing and Publication of Scholarly Work in Medical Journals. That same year, Microbial Cell was selected for inclusion in Clarivate Analytics’ (formerly Thomson Reuters) Emerging Sources Citation Index (ESCI), thus allowing the journal to be accessed through the Web of Science. This selection subsequently allowed for inclusion into additional Web of Science indexes: Biological Abstracts, BIOSIS Previews, Current Contents Life Sciences and Essential Science Indicators. One of the most important milestones was reached in 2017, when Microbial Cell was accepted in Pubmed Central, the archive of biomedical and life sciences journal literature at the U.S. National Institutes of Health’s National Library of Medicine (NIH/NLM). After long evaluation periods, Microbial Cell was further accepted in two of the most selective indexes, Elsevier’s Scopus (2019) and Clarivate Analytics’ Science Citation Index Expanded SCIE (2021).
THE CONCEPT OF MICROBIAL CELL
Over the past few years, Microbial Cell has persistently paid high attention to the quality of its published material. Content-related aspects like novelty, methodology, data presentation, appropriate interpretation, etc., are certainly the main denominators of quality in any submission. In addition, the increasing number of scientific misconduct cases requires special attention. That is why Microbial Cell implements a very strict and careful evaluation of any submitted material in relation to possible data fabrication, data falsification including inadequate manipulation of images and plagiarism. Thus, each submitted work is tested via CrossCheck, a plagiarism detection service powered by the detection software iThenticate. If concerns are raised, Microbial Cell initiates appropriate procedures as detailed by the Committee on Publication Ethics (COPE).
–
With respect to article accessibility, Microbial Cell has always followed an open access approach and used a creative commons (CC) license for copyright purposes. Microbial Cell is published under the CC BY license, which is probably the most generous type of CC licenses. The CC BY license authorizes third parties to share and adapt the published work, even for commercial goals, as long as the authors are appropriately credited and changes are indicated. Thereby, the work can be accessed completely free and with no restrictions; the only prerequisite is the connection to the internet, with no other financial, legal or technical limitations. This approach maximizes the visibility of published work and at the same time, ensures that scientific knowledge is universally and freely accessible to every interested individual around the world. This openness reflects the essential values of science and acts as an effective driver of active research, promoting the free exchange of ideas. Incidentally, 2022 marks the 20th anniversary of the Budapest Open Access Initiative, the public statement of principles relating and defining open access to the research literature.
–
As mentioned, unrestricted article accessibility promotes visibility and increases the impact of a scientific work. The assessment of such an impact involves citations by scholarly journals (because this assessment is peer-connected), which derives in rankings by dedicated indexes. However, we also do acknowledge that alternative outlets, for instance social media, are important indicators of public interest as they increase the diffusion of information at the item (article)-level. Accordingly, Microbial Cell actively uses the corresponding channels including Twitter and Facebook. The journal also provides a social impact measure for each article through PlumX Metrics, a comprehensive monitoring tool that calculates altmetrics for scholarly works.
–
The involvement in, and commitment to, the broad thematic scope of neglected, emerging and trendy microbiology-related topics is a defining characteristic of Microbial Cell. As such, the journal has been continuously supporting the efforts of the microbiology research community well beyond its role as a publication platform. For instance, Microbial Cell runs a waiver program (DevResearch Program) that allows for the partial or complete exemption of article processing charges for corresponding authors based in low-income or lower-middle-income countries. The journal has also sponsored several prizes at and provided support to international conferences, including the Theodor Escherich Symposium on medical microbiome research, the International Symposium “One mitochondrion, many diseases”, or the International Meeting on Yeast Apoptosis.
–
Since the genesis of Microbial Cell, we have been aware of the plethora of facets that make microorganisms a fundamental part of our lives, including at the historical, medical, diagnostic, evolutionary, ecological, environmental, cultural, biotechnological and modelling levels. In accord with this conviction, we have published 100 issues that reflect the wide-ranging importance of microbial research. As we have outlined in this piece, this involves a number of challenges and opportunities that we will continue to embrace in the future.
CONCLUSION
Microbiology has long been at the forefront of research and has attained many achievements in diverse areas of life science and medical practice. Its popularity has gained new heights in recent years, not only due to its ever-increasing applicability, but also due to the unprecedented threat of rising antibiotic and antifungal resistance, as well as due to the re-emergence of old infectious diseases and the emergence of new ones. Indeed, microorganisms represent both opportunity and threat. As the Editors of Microbial Cell, we have the responsibility to ensure adequate selection of publications that represent a high level of contemporary science. At the same time, we have the ambition to provide an open interdisciplinary communication space for microbiologists from all subdisciplines around the globe, in the interest of scientific and societal progress.
REFERENCES
- Dodd MS, Papineau D, Grenne T, Slack JF, Rittner M, Pirajno F, O’Neil J, and Little CTS (2017). Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543(7643): 60–64. 10.1038/nature21377
- Lyons TW, Reinhard CT, and Planavsky NJ (2014). The rise of oxygen in Earth’s early ocean and atmosphere. Nature 506(7488): 307–315. 10.1038/nature13068
- Morens DM, Folkers GK, and Fauci AS (2008). Emerging infections: a perpetual challenge. Lancet Infect Dis 8(11): 710–719. 10.1016/S1473-3099(08)70256-1
- Nathanson N (2016). Chapter 1 – The Human Toll of Viral Diseases: Past Plagues and Pending Pandemics. In: Katze MG, Korth MJ, Law GL, Nathanson N, editors Viral Pathog. Third Ed. Academic Press, Boston; pp 3–16.
- Murray CJ et al. (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 399(10325): 629–655. 10.1016/S0140-6736(21)02724-0
- Rodrigues ML, and Nosanchuk JD (2020). Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl Trop Dis 14(2): e0007964. 10.1371/journal.pntd.0007964
- Perlin DS, Rautemaa-Richardson R, and Alastruey-Izquierdo A (2017). The global problem of antifungal resistance: prevalence, mechanisms, and management. Lancet Infect Dis 17(12): e383–e392. 10.1016/S1473-3099(17)30316-X
- Norman FF, Comeche B, Chamorro S, Pérez-Molina J-A, and López-Vélez R (2020). Update on the major imported protozoan infections in travelers and migrants. Future Microbiol 15(3): 213–225. 10.2217/fmb-2019-0212
- Fletcher SM, Stark D, Harkness J, and Ellis J (2012). Enteric protozoa in the developed world: a public health perspective. Clin Microbiol Rev 25(3): 420–449. 10.1128/CMR.05038-11
- Capela R, Moreira R, and Lopes F (2019). An Overview of Drug Resistance in Protozoal Diseases. Int J Mol Sci 20(22): 5748. 10.3390/ijms20225748
- Peleg AY, Hogan DA, and Mylonakis E (2010). Medically important bacterial–fungal interactions. Nat Rev Microbiol 8(5): 340–349. 10.1038/nrmicro2313
- Vaumourin E, Vourc’h G, Gasqui P, and Vayssier-Taussat M (2015). The importance of multiparasitism: examining the consequences of co-infections for human and animal health. Parasit Vectors 8(1): 545. 10.1186/s13071-015-1167-9
- Salazar F, Bignell E, Brown GD, Cook PC, and Warris A (2022). Pathogenesis of Respiratory Viral and Fungal Coinfections. Clin Microbiol Rev 35(1): e00094-21. 10.1128/CMR.00094-21
- Griffiths EC, Pedersen AB, Fenton A, and Petchey OL (2011). The nature and consequences of coinfection in humans. J Infect 63(3): 200–206. 10.1016/j.jinf.2011.06.005
- Tomley FM, and Shirley MW (2009). Livestock infectious diseases and zoonoses. Philos Trans R Soc B Biol Sci 364(1530): 2637–2642. 10.1098/rstb.2009.0133
- McDonald BA, and Stukenbrock EH (2016). Rapid emergence of pathogens in agro-ecosystems: global threats to agricultural sustainability and food security. Philos Trans R Soc B Biol Sci 371(1709): 20160026. 10.1098/rstb.2016.0026
- Thursby E, and Juge N (2017). Introduction to the human gut microbiota. Biochem J 474(11): 1823–1836. 10.1042/BCJ20160510
- Knight R, Callewaert C, Marotz C, Hyde ER, Debelius JW, McDonald D, and Sogin ML (2017). The Microbiome and Human Biology. Annu Rev Genomics Hum Genet 18: 65–86. 10.1146/annurev-genom-083115-022438
- Cani PD (2018). Human gut microbiome: hopes, threats and promises. Gut 67(9): 1716–1725. 10.1136/gutjnl-2018-316723
- Johnson KV-A, and Foster KR (2018). Why does the microbiome affect behaviour? Nat Rev Microbiol 16(10): 647–655. 10.1038/s41579-018-0014-3
- López-Otín C, and Kroemer G (2021). Hallmarks of health. Cell 184(7): 1929–1939. 10.1016/j.cell.2021.03.033
- Liang G, and Bushman FD (2021). The human virome: assembly, composition and host interactions. Nat Rev Microbiol 19(8): 514–527. 10.1038/s41579-021-00536-5
- Dubey A, Malla MA, Khan F, Chowdhary K, Yadav S, Kumar A, Sharma S, Khare PK, and Khan ML (2019). Soil microbiome: a key player for conservation of soil health under changing climate. Biodivers Conserv 28(8): 2405–2429. 10.1007/s10531-019-01760-5
- Oliver KM, Smith AH, and Russell JA (2014). Defensive symbiosis in the real world – advancing ecological studies of heritable, protective bacteria in aphids and beyond. Funct Ecol 28(2): 341–355. 10.1111/1365-2435.12133
- Roger AJ, Muñoz-Gómez SA, and Kamikawa R (2017). The Origin and Diversification of Mitochondria. Curr Biol CB 27(21): R1177–R1192. 10.1016/j.cub.2017.09.015
- Kroemer G (1997). Mitochondrial implication in apoptosis. Towards an endosymbiont hypothesis of apoptosis evolution. Cell Death Differ 4(6): 443–456. 10.1038/sj.cdd.4400266
- Demain AL (2010). History of Industrial Biotechnology. In: Ind. Biotechnol. John Wiley & Sons, Ltd; pp 17–77.
- Dufossé L, and Fouillaud M (2019). Editorial: Microbial Biotechnology Providing Bio-based Components for the Food Industry. Front Microbiol 10: 2843. 10.3389/fmicb.2019.02843
- Timmusk S, Behers L, Muthoni J, Muraya A, and Aronsson A-C (2017). Perspectives and Challenges of Microbial Application for Crop Improvement. Front Plant Sci 8: 49. 10.3389/fpls.2017.00049
- Shah MP, and Rodriguez-Couto S (2019). Microbial wastewater treatment. Elsevier.
- Gu XY, Wong JWC, and Tyagi RD (2017). 11 – Bioleaching of Heavy Metals From Sewage Sludge for Land Application. In: Wong JW-C, Tyagi RD, Pandey A, editors Curr. Dev. Biotechnol. Bioeng. Elsevier; pp 241–265.
- Head IM, Jones DM, and Röling WFM (2006). Marine microorganisms make a meal of oil. Nat Rev Microbiol 4(3): 173–182. 10.1038/nrmicro1348
- Mehta N, Benzerara K, Kocar BD, and Chapon V (2019). Sequestration of Radionuclides Radium-226 and Strontium-90 by Cyanobacteria Forming Intracellular Calcium Carbonates. Environ Sci Technol 53(21): 12639–12647. 10.1021/acs.est.9b03982
- Tay SB, Natarajan G, Rahim MN bin A, Tan HT, Chung MCM, Ting YP, and Yew WS (2013). Enhancing gold recovery from electronic waste via lixiviant metabolic engineering in Chromobacterium violaceum. Sci Rep 3: 2236. 10.1038/srep02236
- Thompson VS, Gupta M, Jin H, Vahidi E, Yim M, Jindra MA, Nguyen V, Fujita Y, Sutherland JW, Jiao Y, and Reed DW (2018). Techno-economic and Life Cycle Analysis for Bioleaching Rare-Earth Elements from Waste Materials. ACS Sustain Chem Eng 6(2): 1602–1609. 10.1021/acssuschemeng.7b02771
- Cockell CS et al. (2020). Space station biomining experiment demonstrates rare earth element extraction in microgravity and Mars gravity. Nat Commun 11(1): 5523. 10.1038/s41467-020-19276-w
- Keasling J, Garcia Martin H, Lee TS, Mukhopadhyay A, Singer SW, and Sundstrom E (2021). Microbial production of advanced biofuels. Nat Rev Microbiol 19(11): 701–715. 10.1038/s41579-021-00577-w
- Asensio Y, Mansilla E, Fernandez-Marchante CM, Lobato J, Cañizares P, and Rodrigo MA (2017). Towards the scale-up of bioelectrogenic technology: stacking microbial fuel cells to produce larger amounts of electricity. J Appl Electrochem 47(10): 1115–1125. 10.1007/s10800-017-1101-2
- Pham JV, Yilma MA, Feliz A, Majid MT, Maffetone N, Walker JR, Kim E, Cho HJ, Reynolds JM, Song MC, Park SR, and Yoon YJ (2019). A Review of the Microbial Production of Bioactive Natural Products and Biologics. Front Microbiol 10: 1404. 10.3389/fmicb.2019.01404
- Bustamante-Torres M, Romero-Fierro D, Estrella-Nuñez J, Cuadros-Buenaventura E, and Bucio E (2022). Application of Microbes in Biotechnology, Industry, and Medical Field. In: Inamuddin, Ahamed MI, Prasad R, editors Appl. Microbes Environ. Microb. Biotechnol. Springer, Singapore; pp 113–152.
- Blount ZD (2015). The unexhausted potential of E. coli. eLife 4: e05826. 10.7554/eLife.05826
- Leigh JA, Albers S-V, Atomi H, and Allers T (2011). Model organisms for genetics in the domain Archaea: methanogens, halophiles, Thermococcales and Sulfolobales. FEMS Microbiol Rev 35(4): 577–608. 10.1111/j.1574-6976.2011.00265.x
- Errington J, and van der Aart LT (2020). Microbe Profile: Bacillus subtilis: model organism for cellular development, and industrial workhorse. Microbiology 166(5): 425–427. 10.1099/mic.0.000922
- Hernández-Prieto MA, Semeniuk TA, Giner-Lamia J, and Futschik ME (2016). The Transcriptional Landscape of the Photosynthetic Model Cyanobacterium Synechocystis sp. PCC6803. Sci Rep 6(1): 22168. 10.1038/srep22168
- Govers SK, and Jacobs-Wagner C (2020). Caulobacter crescentus: model system extraordinaire. Curr Biol CB 30(19): R1151–R1158. 10.1016/j.cub.2020.07.033
- Harris E (2008). The Chlamydomonas Sourcebook: Introduction to Chlamydomonas and Its Laboratory Use, 2nd edition. Academic Press.
- Matt G, and Umen J (2016). Volvox: A simple algal model for embryogenesis, morphogenesis and cellular differentiation. Dev Biol 419(1): 99–113. 10.1016/j.ydbio.2016.07.014
- Marshall WF (2021). Regeneration in Stentor coeruleus. Front Cell Dev Biol 9: 753625. 10.3389/fcell.2021.753625
- Russell JJ, Theriot JA, Sood P, Marshall WF, Landweber LF, Fritz-Laylin L, Polka JK, Oliferenko S, Gerbich T, Gladfelter A, Umen J, Bezanilla M, Lancaster MA, He S, Gibson MC, Goldstein B, Tanaka EM, Hu C-K, and Brunet A (2017). Non-model model organisms. BMC Biol 15(1): 55. 10.1186/s12915-017-0391-5
- Bolotin-Fukuhara M, Dumas B, and Gaillardin C (2010). Yeasts as a model for human diseases. FEMS Yeast Res 10(8): 959–960. 10.1111/j.1567-1364.2010.00693.x
- Ruetenik A, and Barrientos A (2018). Exploiting Post-mitotic Yeast Cultures to Model Neurodegeneration. Front Mol Neurosci 11: 400. 10.3389/fnmol.2018.00400
- Ferreira R, Limeta A, and Nielsen J (2019). Tackling Cancer with Yeast-Based Technologies. Trends Biotechnol 37(6): 592–603. 10.1016/j.tibtech.2018.11.013
- Madeo F, Fröhlich E, and Fröhlich KU (1997). A yeast mutant showing diagnostic markers of early and late apoptosis. J Cell Biol 139(3): 729–734. 9348289
- Madeo F, Fröhlich E, Ligr M, Grey M, Sigrist SJ, Wolf DH, and Fröhlich KU (1999). Oxygen stress: a regulator of apoptosis in yeast. J Cell Biol 145(4): 757–767. 10.1083/jcb.145.4.757
- Carmona-Gutierrez D et al. (2018). Guidelines and recommendations on yeast cell death nomenclature. Microb Cell 5(1): 4–31. 10.15698/mic2018.01.607
- Carmona-Gutierrez D et al. (2019). The flavonoid 4,4′-dimethoxychalcone promotes autophagy-dependent longevity across species. Nat Commun 10(1): 651. 10.1038/s41467-019-08555-w
- Ohsumi Y (2014). Historical landmarks of autophagy research. Cell Res 24(1): 9–23. 10.1038/cr.2013.169
- Longo VD, Shadel GS, Kaeberlein M, and Kennedy B (2012). Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab 16(1): 18–31. 10.1016/j.cmet.2012.06.002
- Feyder S, De Craene J-O, Bär S, Bertazzi DL, and Friant S (2015). Membrane Trafficking in the Yeast Saccharomyces cerevisiae Model. Int J Mol Sci 16(1): 1509–1525. 10.3390/ijms16011509
- Novick P, Ferro S, and Schekman R (1981). Order of events in the yeast secretory pathway. Cell 25(2): 461–469. 10.1016/0092-8674(81)90064-7
- Mentel M, Chovančíková P, Zeman I, and Polčic P (2021). Learning from Yeast about Mitochondrial Carriers. Microorganisms 9(10): 2044. 10.3390/microorganisms9102044
- Neupert W, and Herrmann JM (2007). Translocation of proteins into mitochondria. Annu Rev Biochem 76: 723–749. 10.1146/annurev.biochem.76.052705.163409
- Chacinska A, Koehler CM, Milenkovic D, Lithgow T, and Pfanner N (2009). Importing mitochondrial proteins: machineries and mechanisms. Cell 138(4): 628–644. 10.1016/j.cell.2009.08.005
- Hartwell LH, Culotti J, Pringle JR, and Reid BJ (1974). Genetic control of the cell division cycle in yeast. Science 183(4120): 46–51. 10.1126/science.183.4120.46
- Eisenberg T, Knauer H, Schauer A, Büttner S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes C, Antonacci L, Fussi H, Deszcz L, Hartl R, Schraml E, Criollo A, Megalou E, Weiskopf D, Laun P, Heeren G, Breitenbach M, Grubeck-Loebenstein B, Herker E, Fahrenkrog B, Fröhlich K-U, Sinner F, Tavernarakis N, Minois N, Kroemer G, and Madeo F (2009). Induction of autophagy by spermidine promotes longevity. Nat Cell Biol 11(11): 1305–1314. 10.1038/ncb1975
- Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, and Sinclair DA (2006). Resveratrol improves health and survival of mice on a high-calorie diet. Nature 444(7117): 337–342. 10.1038/nature05354
- Zimmermann A, Hofer S, Pendl T, Kainz K, Madeo F, and Carmona-Gutierrez D (2018). Yeast as a tool to identify anti-aging compounds. FEMS Yeast Res 18(6): foy020. 10.1093/femsyr/foy020
- Menacho-Márquez M, and Murguía JR (2007). Yeast on drugs: Saccharomyces cerevisiae as a tool for anticancer drug research. Clin Transl Oncol Off Publ Fed Span Oncol Soc Natl Cancer Inst Mex 9(4): 221–228. 10.1007/s12094-007-0043-2
- Tebbets B, Stewart D, Lawry S, Nett J, Nantel A, Andes D, and Klein BS (2012). Identification and Characterization of Antifungal Compounds Using a Saccharomyces cerevisiae Reporter Bioassay. PLOS ONE 7(5): e36021. 10.1371/journal.pone.0036021
- Bilsland E, Sparkes A, Williams K, Moss HJ, de Clare M, Pir P, Rowland J, Aubrey W, Pateman R, Young M, Carrington M, King RD, and Oliver SG Yeast-based automated high-throughput screens to identify anti-parasitic lead compounds. Open Biol 3(2): 120158. 10.1098/rsob.120158
- Duina AA, Miller ME, and Keeney JB (2014). Budding Yeast for Budding Geneticists: A Primer on the Saccharomyces cerevisiae Model System. Genetics 197(1): 33–48. 10.1534/genetics.114.163188
- Hoffman CS, Wood V, and Fantes PA (2015). An Ancient Yeast for Young Geneticists: A Primer on the Schizosaccharomyces pombe Model System. Genetics 201(2): 403–423. 10.1534/genetics.115.181503
- Nurse P, Thuriaux P, and Nasmyth K (1976). Genetic control of the cell division cycle in the fission yeast Schizosaccharomyces pombe. Mol Gen Genet MGG 146(2): 167–178. 10.1007/BF00268085
- Carmona-Gutierrez D, Kroemer G, and Madeo F (2014). One cell, one love: a journal for microbial research. Microbial Cell 1(1): 1–5. 10.15698/mic2014.01.118
- Piatkov KI, Vu TTM, Hwang C-S, and Varshavsky A (2015). Formyl-methionine as a degradation signal at the N-termini of bacterial proteins. Microbial Cell 2(10): 376–393. 10.15698/mic2015.10.231
- Bachmair A, Finley D, and Varshavsky A (1986). In vivo half-life of a protein is a function of its amino-terminal residue. Science 234(4773): 179–186. 10.1126/science.3018930
- Varshavsky A (2006). The early history of the ubiquitin field. Protein Sci Publ Protein Soc 15(3): 647–654. 10.1110/ps.052012306
- Yin Z, Pascual C, and Klionsky DJ (2016). Autophagy: machinery and regulation. Microbial Cell 3(12): 588–596. 10.15698/mic2016.12.546
- Takeshige K, Baba M, Tsuboi S, Noda T, and Ohsumi Y (1992). Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J Cell Biol 119(2): 301–311. 10.1083/jcb.119.2.301
- Tsukada M, and Ohsumi Y (1993). Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett 333(1–2): 169–174. 10.1016/0014-5793(93)80398-e
- Tooze SA, and Dikic I (2016). Autophagy Captures the Nobel Prize. Cell 167(6): 1433–1435. 10.1016/j.cell.2016.11.023
- García-Martínez J, Maldonado RD, Guzmán NM, and Mojica FJM (2018). The CRISPR conundrum: evolve and maybe die, or survive and risk stagnation. Microbial Cell 5(6): 262–268. 10.15698/mic2018.06.634
- Mojica FJ, Juez G, and Rodríguez-Valera F (1993). Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol Microbiol 9(3): 613–621. 10.1111/j.1365-2958.1993.tb01721.x
- Mojica FJM, Díez-Villaseñor C, García-Martínez J, and Soria E (2005). Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J Mol Evol 60(2): 174–182. 10.1007/s00239-004-0046-3
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, and Charpentier E (2012). A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 337(6096): 816–821. 10.1126/science.1225829
- Microbial Cell Special Issue repositorium. http://microbialcell.com/special-issues/. [Accessed 11.03.2022]
–
COPYRIGHT
© 2022

A hundred spotlights on microbiology: how microorganisms shape our lives by Carmona-Gutierrez et al. is licensed under a Creative Commons Attribution 4.0 International License.








