
Viewpoint:
Microbial Cell, Vol. 7, No. 5, pp. 119 - 128; doi: 10.15698/mic2020.05.715
Digesting the crisis: autophagy and coronaviruses

1 Institute for Molecular Biosciences, NAWI Graz, University of Graz, Graz, Austria.
2 BioHealth Graz, Graz, Austria.
3 Centre de Recherche des Cordeliers, Equipe labellisée par la Ligue contre le cancer, Université de Paris, Sorbonne Université, INSERM U1138, Institut Universitaire de France, Paris, France.
4 Metabolomics and Cell Biology Platforms, Institut Gustave Roussy, Villejuif, France.
5 Pôle de Biologie, Hôpital Européen Georges Pompidou, AP-HP, Paris, France.
6 Suzhou Institute for Systems Medicine, Chinese Academy of Medical Sciences, Suzhou, China.
7 Karolinska Institute, Department of Women’s and Children’s Health, Karolinska University Hospital, Stockholm, Sweden.
8 BioTechMed Graz, Graz, Austria.
Keywords: COVID-19, SARS, SARS-CoV-2, MERS, coronavirus, virophagy, inflammation, immunity.
Abbreviations:
CoV – coronavirus;
COVID-19 – Coronavirus Disease 2019;
DMV – double membrane vesicle;
hCoV – human pathogenic CoV;
HIV – human immunodeficiency virus;
HSV – herpes simplex virus;
LIR – LC3-interacting region;
MERS – Middle East respiratory syndrome;
MHC – major histocompatibility complex;
MHV – mouse hepatitis virus;
PAMP – pathogen-associated molecular pattern;
PEDV – porcine epidemic diarrhea virus;
PRR – pattern recognition receptor;
SARS – severe acute respiratory syndrome;
TGEV – transmissible gastroenteritis virus;
UBD – ubiquitin-binding domain.
Received originally: 15/04/2020 Received in revised form: 30/05/2020
Accepted: 01/05/2020
Published: 04/05/2020
Correspondence:
Didac Carmona-Gutierrez, Institute of Molecular Biosciences, Humboldtstrasse 50/EG, 8010 - Graz, Austria; carmonag@uni-graz.at
Frank Madeo, Institute of Molecular Biosciences, Humboldtstrasse 50/EG, 8010 - Graz, Austria; frank.madeo@uni-graz.at
Conflict of interest statement: D.C-G., G.K. and F.M. are the scientific co-founders of Samsara Therapeutics, a company that develops novel pharmacological autophagy inducers. A.Z. has equity interests in Samsara Therapeutics. F.M. and D.C-G. have equity interests in TLL (The Longevity Labs), a company that develops natural food extracts.
Please cite this article as: Didac Carmona-Gutierrez, Maria A. Bauer, Andreas Zimmermann, Katharina Kainz, Sebastian J. Hofer, Guido Kroemer and Frank Madeo (2020). Digesting the crisis: autophagy and coro-naviruses. Microbial Cell 7(5): 119-128. doi: 10.15698/mic2020.05.715
Abstract
Autophagy is a catabolic pathway with multifaceted roles in cellular homeostasis. This process is also involved in the antiviral response at multiple levels, including the direct elimination of intruding viruses (virophagy), the presentation of viral antigens, the fitness of immune cells, and the inhibition of excessive inflammatory reactions. In line with its central role in immunity, viruses have evolved mechanisms to interfere with or to evade the autophagic process, and in some cases, even to harness autophagy or constituents of the autophagic machinery for their replication. Given the devastating consequences of the current COVID-19 pandemic, the question arises whether manipulating autophagy might be an expedient approach to fight the novel coronavirus SARS-CoV-2. In this piece, we provide a short overview of the evidence linking autophagy to coronaviruses and discuss whether such links may provide actionable targets for therapeutic interventions.
INTRODUCTION
The ongoing outbreak of the respiratory disease COVID-19 (Coronavirus Disease 2019) is endangering individuals, governments and societies around the world. Indeed, this pandemic has unprecedented global consequences at the economic and social levels as it challenges the medical and research communities. The first case of COVID-19 was reported in December 2019 in Wuhan (China) [1] and, despite increasingly drastic efforts to contain the disease, the infection rapidly spread across the planet, leading the WHO to declare it, first, a global health emergency (in January 2020) and eventually a pandemic (on March 11, 2020) [2][3]. As of April 30th, more than 200 countries and territories have reported COVID-19 cases with a total of over three million confirmed cases and more than 220.000 confirmed deaths [4]. In the current absence of effective treatments and vaccines, these numbers will continue to grow inexorably. The main clinical features of COVID-19 (fever, dry cough, pneumonia) manifest after an average incubation period of approximately 5 days [5][6] with severe cases advancing towards severe acute respiratory syndrome (SARS) [7][8][9]. COVID-19 was quickly shown to be caused by a novel coronavirus (CoV) that was named SARS-CoV-2 [7][10].
–
SARS-CoV-2 is structurally related to the members of the Coronaviridae family in the order Nidovirales[10][11]. CoVs are enveloped viruses with a single-strand, positive-sense RNA genome and are separated into four genera according to their genome characteristics: α-CoV, β-CoV, γ-CoV, and δ-CoV [12][13]. Until the emergence of COVID-19, six human pathogenic CoVs (hCoVs) had been identified: four non-SARS-like hCoVs (HCoV 229E, NL63, OC43, and HKU1), which do not cause major (fatal) epidemics, and two SARS-like hCoVs (SARS-CoV and Middle East respiratory syndrome coronavirus MERS-CoV) [14]. Both SARS-like hCoVs, which belong to the β-CoV genus, have already caused major outbreaks and health emergencies in China (SARS-CoV, 2002-2003) as well as in the Middle East (MERS-CoV, 2012) [15]. The novel SARS-CoV-2 increases the number of known hCoVs to seven, all of which have a zoonotic origin, meaning that they originate from vertebrate animals (possibly bats for SARS-CoV-2) and have reached humans via interspecies transmission [16].
AUTOPHAGY AS A PROTECTIVE CATABOLIC PROCESS
Given the devastating scenario that COVID-19 has already generated, a multitude of potential approaches against SARS-CoV-2 are currently being conceived. In that sense, we think that the manipulation of autophagy might be worth exploring. Macroautophagy (hereafter referring to autophagy) is an intracellular catabolic mechanism that regulates the elimination of unwanted, superfluous or defective cell components [17]. In this process, the targeted molecules/organelles are enclosed in double-membrane vesicles called autophagosomes, which eventually fuse with lysosomes for degradation of the delivered contents. This process is induced upon energy deprivation and/or physical activity in order to secure sufficient nutrient supply and is thus often referred to as a cellular self-digestion system. The compounds resulting from lysosomal degradation (e.g., fatty acids, amino acids) may be released into the cytoplasm for the generation of new macromolecules and/or bioenergetics reactions. Autophagy, therefore, represents a recycling mechanism under circumstances of low resource availability [17].
–
The digestion of damaged and thus potentially detrimental cargo also increases the fitness at the cellular and consequently at the organismal level. Accordingly, the induction of autophagy by dietary interventions (e.g., caloric restriction, fasting regimens), behavioral cues (e.g., exercise) or pharmacological agents (e.g., caloric restriction mimetics like spermidine, rapamycin, resveratrol or specific chalcones [18][19][20][21][22]) has been consistently linked to beneficial effects on health [23]. In turn, reduced autophagic capacity is connected to aging progression and a number of pathologies including cardiovascular disease, cancer and neurogeneration [24].
–
The autophagic machinery can retrieve its cargo in bulk, but it can also selectively target distinct intracellular components and organelles via specific adapter proteins that interact with both the individual substrates and the autophagosomes [25]. Specifically, these autophagy adaptors (e.g., p62/SQSTM1, optineurin) contain both a ubiquitin-binding domain (UBD) and an LC3-interacting region (LIR). The UBD recognizes ubiquitin tags decorating the specific target, while the LIR allows binding of LC3 proteins attached to nascent phagophores, the structures that eventually close to form autophagosomes [26]. Accordingly, autophagic subforms have been defined that describe, for example, the specific removal of damaged mitochondria (mitophagy), the disposal of protein aggregates (aggrephagy), the digestion of lipid droplets (lipophagy), etc [27].
–
Yet another selective autophagy subroutine is xenophagy, the specialized elimination of intracellular pathogens, including fungi, bacteria and viruses [28]. The xenophagic disposal of viruses (sometimes also termed virophagy) has been described for different viral pathogens, including human immunodeficiency virus-1 (HIV-1) [29][30] and herpes simplex virus-1 (HSV-1) [31][32]. However, many viruses (including HIV-1 and HSV-1) and other pathogens have also evolved strategies to escape or inhibit autophagy and sometimes even to manipulate the autophagic machinery for their replicative benefit [33]. Thus, autophagy emerges as a crucial element in the evolutionary-driven arms race of host against pathogen. This further underlines the significance of xenophagy as a central antimicrobial mechanism, but also limits the potential of autophagy induction to fight infections.
CORONAVIRUSES AND AUTOPHAGY
The question whether autophagy induction might be beneficial to specifically counteract SARS-CoV-2 infection cannot be answered at this point. However, the existing data on other CoVs suggest that – despite some conflicting results – autophagy induction might be a valid approach that should be subjected to further evaluation. The autophagy-lysosomal system does indeed seem to play a central role during the infection with different CoVs, including SARS-CoVs [34][35][36][37]. Nevertheless, the diverse putative roles of autophagy during viral infection [38] require that two opposite aspects be covered by this overview: (i) Is there evidence for autophagy being used for the survival or replication of CoVs (proviral effects)? (ii) Can autophagy induction reduce cellular infection, and does the virus actively inhibit autophagy (suggesting antiviral effects for autophagy)? Of note, cellular manipulation of autophagic levels during infection may also reflect desperate attempts of the cell to reestablish homeostasis, either through restriction of viral entry by actively shunting endocytosis/endosomal trafficking (possibly resulting in autophagy reduction as a side-effect) [39] or to counteract virally induced cell death by increasing cytoprotective autophagy. As a third major aspect, (iii) the autophagic effects on antiviral immunity must be taken into consideration. Over the last 15 years, a number of studies have addressed these issues in the context of the “old” CoVs, those that historically precede SARS-CoV-2.
–
Can autophagy be highjacked for CoV replication?
CoVs rely on the formation of replication complexes at double membrane vesicles (DMVs), where viral genome replication and transcription occurs [40][41]. As it may be at least partly the case for autophagosomes, DMVs probably depend on ER-derived membranes for their generation [40][42]. Therefore, it was initially suspected that CoVs might usurp the autophagosomal machinery for DMV formation. In fact, an early study using ATG5 knockout cells implied that autophagy may promote mouse hepatitis virus (MHV) replication via formation of DMVs [43]. In a follow-up work, the same authors reported the co-localization of specific replication proteins (nsp2, nsp3, nsp8) with endogenous LC3, suggesting a connection between replication and autophagosome formation [44]. Other studies reported that the porcine CoVs transmissible gastroenteritis virus (TGEV) [45] and porcine epidemic diarrhea virus (PEDV) [46] may induce autophagy to promote replication. However, the link between autophagy and replication of CoVs has been challenged by a number of different reports, showing that deletion of the essential autophagy genes ATG5 or ATG7 does not affect the replication of MHV [47][48] or SARS-CoV [49]. Furthermore, the co-localization of LC3 (or GFP-LC3) with the SARS-CoV RNA replication complex could not be confirmed [50]. A study using pharmacological induction (rapamycin) or genetic inhibition (knockdown of LC3, ATG5, or ATG7) of autophagy even demonstrated that TGEV replication is negatively regulated by autophagy [51]. In addition, rapamycin reduced infectivity of PEDV [52]. Altogether, although some conflicting results exist, autophagy may not be primarily engaged in promoting CoV replication.
–
However, single components of the autophagic machinery may be hijacked independently of their activity in autophagic processing. Non-lipidated LC3, for instance, is used for membrane derivation for DMVs in CoVs, and downregulation of LC3 but not inactivation of autophagy, counteracts CoV infection [47]. Notably, some components of the canonical autophagy machinery can target invading pathogens by promoting phagosome-to-lysosome fusion [53]. During LC3-associated phagocytosis (LAP), for instance, which does not involve sequestration in autophagosomes, LC3 is recruited to phagosome membranes to facilitate lysosomal fusion and degradation [54]. In addition, it has been proposed that non-canonical autophagy and intruding viruses, both of which can be inhibited by the fungal compound brefeldin A, may converge in the retrieval of membranes [55]. Non-canonical autophagy includes ATG5- and ATG7-independent as well as Beclin-1-independent autophagy [56][57]. Altogether, these observations add further layers of possible modulation of autophagic processes in trying to attenuate viral infection (Figure 1).
–
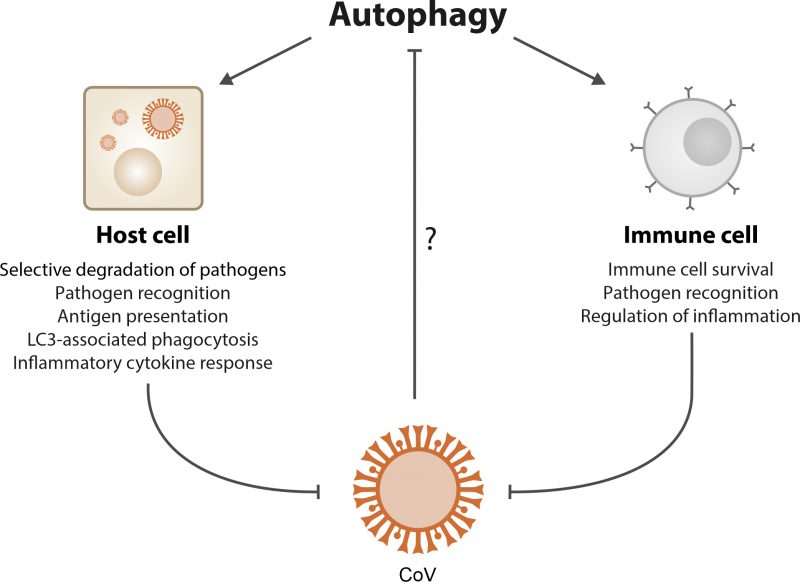 | FIGURE 1: Possible protective mechanisms of autophagy against coronaviruses. Autophagy may counteract viral infection by executing and supporting different antiviral pathways in infected host cells as well as by promoting the function and survival of immune cells. Therefore, an increase in autophagic flux may be effective against coronaviruses (CoVs). Accordingly, CoVs may have developed strategies to limit cellular and organismal immunity by preventing autophagy. |
–
Does autophagy counteract CoV infection?
Intriguingly, a body of evidence indicates that CoVs may actually inhibit autophagy, which in turn teleologically suggests an antiviral role for autophagy. Accordingly, a number of studies have shown that increasing autophagic capacity may be beneficial against infection (Figure 1). A temporal kinome analysis of MERS-CoV-infected hepatocytes demonstrated selective activation of the ERK/MAPK and PI3K/AKT/mTOR signaling responses, both of which have inhibitory effects on autophagy [58]. Accordingly, pharmacological inhibition of these pathways was able to inhibit MERS-CoV infection [58]. Moreover, although one of the central replicase proteins (nsp6) of avian infectious bronchitis virus (IBV) seems to promote autophagosome formation at the omegasome level [59], nsp6 also limits further autophagosomal expansion [60], thus compromising autophagic delivery of viral components to lysosomes. Interestingly, nsp6 is also present in other CoVs, including SARS-CoV-2 [61].
–
Overexpression of the protease PLP2 of SARS-CoV and MERS-CoV in different cell lines inhibited autophagosome-lysosome fusion and autophagic flux [62]. A recent report corroborated the autophagy-inhibitory activity of MERS-CoV and showed that induction of autophagy can reduce MERS-CoV replication [37]. Specifically, the authors found that pharmacological inhibition of the ubiquitin ligase SKP2 increases the levels of Beclin-1, a central regulator of phagophore formation [37]. SKP2 executes lysine-48-linked poly-ubiquitination of Beclin-1, targeting it for proteasomal degradation, the inhibition of which promoted autophagy and efficiently reduced MERS-CoV replication. This work also shows that ectopic expression of at least three viral proteins (nsp6, p4b, p5) restricts autophagy [37]. While nsp6 may limit autophagosome expansion (see above), p4b inhibits RNAse L activation (a pro-autophagic event [63]), and p5 has been connected to the blockage of IFN-β induction [63][64], which itself may be linked to autophagy [65]. Accordingly, deletion of p4b or p5 resulted in reduced MERS-CoV growth, although disputing some previous observations, which the authors explain by methodological differences [37]. Thus, the group-specific accessory proteins, which by definition are not essential for viral replication but are involved in the modulation of host cells and immune evasion [66][67], may represent targets for reducing the autophagy-inhibitory effects of CoVs.
–
The FDA-approved anti-malarial drugs chloroquine and hydroxychloroquine have been suggested to be repurposed for the treatment of COVID-19 [68][69][70], but this remains widely controversial [71][72][73]. Although chloroquine is a lysosomotropic agent that blocks autophagic degradation, possibly by impairing autophagosome fusion with lysosomes [74], the putative effects on autophagy may not be necessarily causal for the antiviral activity. In fact, endosomal acidification after endocytosis is critical for SARS-CoV-2 entry [75], and chloroquine inhibits this acidification [76]. In addition, chloroquine limits terminal glycosylation of the metallopeptidase ACE2, the functional receptor for SARS-CoV and SARS-CoV-2 cell entry [68][75][77]. Non-glycosylated ACE2 seems to interact less efficiently with the SARS-CoV spike protein, thus reducing viral entry [78]. These modes of action would target the virus upstream of autophagy, making it unlikely that autophagy modulation contributes to the outcome of chloroquine treatment at that point. Additionally, chloroquine has been shown to induce autophagy-independent effects, for instance, Golgi disorganization [74] and pulmonary vasodilation [79] that may contribute to its controversial clinical activity. Interestingly, the net effects on autophagy may differ depending on parameters like cell type and dose: for instance, chloroquine-mediated lysosomal stress may promote the nuclear translocation of the pro-autophagic transcription factors TFEB and TFE3 [80][81]. Altogether, further clinical work must clarify if chloroquine and hydroxychloroquine have major effects on COVID-19 and at which (early?) phase of the disease these drugs should be administered. If chloroquine or its derivative turned out to be clinically useful against COVID-19, it will be important to understand to which extent such effects are connected to the modulation of autophagy.
–
Indeed, repurposing of known (especially currently FDA-approved) drugs might offer a welcome shortcut to rapidly developing treatments against COVID-19. Obviously, even in the present period of desperate search for pharmacological treatments, the efficacy (and lack of side-effects) of anti-COVID-19 treatments needs to be proven by clinical studies subjected to rigorous scientific scrutiny. While data regarding such attempts remain preliminary at this stage, several currently available preprints allow the speculation of autophagy as a possible druggable target. For instance, a recent preprint mapping the SARS-CoV-2 interactome in human host cells identifies several host factors connected to autophagy [82]. Amongst others, the authors find viral-human interactions with proteins directly modulated by mammalian target of rapamycin complex 1 (mTORC1) [82], a master regulator of cell proliferation and autophagy known to be affected by other viruses [83][84].
–
Intriguingly, another recent preprint presents in vitro data showing that SARS-CoV-2 infection restricts autophagy and that, in turn, pro-autophagic compounds – including spermidine – may inhibit viral propagation [85]. Admittedly, spermidine has a plethora of physiological functions, including regulation of RNA-to-protein translation, as well as amelioration of RNA and DNA stability [86], and these effects need further investigation, specifically in relation with a possible autophagy induction. In fact, the role of spermidine during infection may be pleiotropic. On the one hand, spermidine is an autophagy inducer [18][87][88][89][90] and may promote viral xenophagy; also, it may contribute to endosomal pH buffering [91] and thus block viral entry. On the other hand, polyamines are essential for viral replication [92], and the overexpression of spermidine/spermine-N(1)-acetyltransferase by the host, which drives spermidine (and spermine) depletion, is a common cellular response to viral infections, including that of RNA viruses [93]. This underlines once more that any pharmacological candidate for the fight against SARS-CoV-2 should be examined for its (possible) pleiotropy during infection.
–
The role of autophagy on immune signaling
Autophagy directly impacts immune signaling, and autophagy induction hence may counteract CoVs by promoting immunological defense mechanisms (Figure 1). For instance, autophagy regulates, and can be regulated by, so-called pattern recognition receptors (PRRs), which sense molecules specifically derived from microorganisms (pathogen-associated molecular patterns, PAMPs) [94][95]. PRRs are present in various cells of the innate immune system and many epithelial cells [95]. There, they are distributed at diverse locations (either at the cell surface or in the cytoplasm), providing a recognition system for different life cycle phases of a given pathogen [95]. PAMP-induced signaling triggers a multitude of pathways, including pro-inflammatory responses, that result in the synthesis of immunostimulatory molecules including cytokines, chemokines and immunoreceptors [94].
–
Autophagy further assists immune responses by processing and supporting the presentation of antigens on major histocompatibility complex (MHC) class II, responsible for loading extracellular antigens [96][97]. In addition, autophagy can deliver exogenous viral antigens into MHC-I molecules for cross-presentation (the MHC-I pathway is usually employed for loading endogenous antigens) [97][98]. Moreover, autophagy directly impacts the activation, proliferation and differentiation of immune cells [99]. Amongst others, autophagy has been connected to the differentiation of CD8+ T-cells into cytotoxic T lymphocytes, to dendritic cell and B cell development, to plasma cell differentiation, and to macrophage differentiation, therefore covering multiple instances of the innate and adaptive immune systems [99]. Of note, autophagy activation by caloric restriction mimetics (including hydroxycitric acid and spermidine) also improves anticancer immunosurveillance [100]. In addition, the general cellular pro-survival effects of autophagy extend to several immune cell types. Pharmacological autophagy induction by spermidine [87], for instance, has been demonstrated to promote the maintenance and survival of memory CD8+ T cells [101] as well as to counteract B lymphocyte senescence [102].
–
The interplay of autophagy and inflammation is complex, since it does not only encompass positive but also negative regulatory mechanisms. For example, autophagy can digest the interleukin precursors produced by inflammasomes (e.g. pro-IL-1), but also directly target inflammasome components (e.g. NLRP3, AIM2 and ASC) [103]. In sum, autophagy ensures acute inflammatory responses while preventing excessive and prolonged hyperinflammation.
–
Finally, it should be noted that autophagy may have paracrine functions in the form of unconventional secretion [104], thus adding yet another layer of non-cell autonomous effects. In the context of viral infections, this mechanism may be used to induce protective responses in cells neighboring an infection site [105][106]. In primary human placental trophoblasts, for instance, autophagy induction promotes packaging and exosome-dependent delivery of specific miRNAs, which induce autophagy in non-placental recipient cells, conferring resistance to a variety of viruses [105]. This may be (one of) the mechanism(s) through which placental and maternal cells optimize their defense against viral infections during pregnancy.
CONCLUSIONS
The existing evidence allows the speculation that autophagy induction might counteract CoV infection at different levels, although more specific data are certainly required. As mentioned above, the restriction of calorie intake is the most robust method to induce autophagy. However, the fight against acute infections also requires sufficient energy supply, suggesting that autophagy induction via caloric restriction or fasting regimens may be counterproductive, at least in the short-term during (as well as shortly before) infection. Therefore, caloric restriction mimetics [107][108], natural or synthetic compounds with the ability to induce autophagy, may circumvent this problem.
–
Irrespectively of whether autophagy modulation will eventually be part of the strategies against COVID-19, the current pandemic outbreak is a shocking reminder that emerging (and re-emerging) infectious pathogens are (and will be) a major challenge. In view of the exposed vulnerability of our medical structures and socioeconomic well-being, this pandemic underlines how essential it is to further establish and secure global healthcare as well as to promote and extend robust research against infectious diseases.
REFERENCES
- Yi Y, Lagniton PNP, Ye S, Li E, and Xu R-H (2020). COVID-19: what has been learned and to be learned about the novel coronavirus disease. Int J Biol Sci 16(10): 1753–1766. doi: 10.7150/ijbs.45134
- Rismanbaf A (2020). Potential Treatments for COVID-19; a Narrative Literature Review. Arch Acad Emerg Med 8(1): e29. PMID: 32232214
- Zhou P, Yang X-L, Wang X-G, Hu B, Zhang L, Zhang W, Si H-R, Zhu Y, Li B, Huang C-L, Chen H-D, Chen J, Luo Y, Guo H, Jiang R-D, Liu M-Q, Chen Y, Shen X-R, Wang X, Zheng X-S, Zhao K, Chen Q-J, Deng F, Liu L-L, Yan B, Zhan F-X, Wang Y-Y, Xiao G-F, and Shi Z-L (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579(7798): 270–273. doi: 10.1038/s41586-020-2012-7
- World Health Organization (2020). Coronavirus. Available at: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 [Accessed: 30.04.2020]
- Guan W-J et al. (2020). Clinical Characteristics of Coronavirus Disease 2019 in China. N Engl J Med. doi: 10.1056/NEJMoa2002032
- Lauer SA, Grantz KH, Bi Q, Jones FK, Zheng Q, Meredith HR, Azman AS, Reich NG, and Lessler J (2020). The Incubation Period of Coronavirus Disease 2019 (COVID-19) From Publicly Reported Confirmed Cases: Estimation and Application. Ann Intern Med10.7326/M20-0504
- Gorbalenya AE, Baker SC, Baric RS, Groot RJ de, Drosten C, Gulyaeva AA, Haagmans BL, Lauber C, Leontovich AM, Neuman BW, Penzar D, Perlman S, Poon LLM, Samborskiy D, Sidorov IA, Sola I, and Ziebuhr J (2020). The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol 5: 536–544. doi: 10.1038/s41564-020-0695-z
- Chen N, Zhou M, Dong X, Qu J, Gong F, Han Y, Qiu Y, Wang J, Liu Y, Wei Y, Xia J, Yu T, Zhang X, and Zhang L (2020). Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet Lond Engl 395(10223): 507–513. doi: 10.1016/S0140-6736(20)30211-7
- Ren L-L et al. (2020). Identification of a novel coronavirus causing severe pneumonia in human: a descriptive study. Chin Med J (Engl). doi: 10.1097/CM9.0000000000000722
- Velavan TP, and Meyer CG (2020). The COVID-19 epidemic. Trop Med Int Health 25(3): 278–280. doi: 10.1111/tmi.13383
- Schoeman D, and Fielding BC (2019). Coronavirus envelope protein: current knowledge. Virol J 16(1): 69. doi: 10.1186/s12985-019-1182-0
- Forni D, Cagliani R, Clerici M, and Sironi M (2017). Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol 25(1): 35–48. doi: 10.1016/j.tim.2016.09.001
- Su S, Wong G, Shi W, Liu J, Lai ACK, Zhou J, Liu W, Bi Y, and Gao GF (2016). Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol 24(6): 490–502. doi: 10.1016/j.tim.2016.03.003
- Raoult D, Zumla A, Locatelli F, Ippolito G, and Kroemer G (2020). Coronavirus infections: Epidemiological, clinical and immunological features and hypotheses. Cell Stress 4(4):66-75. doi: 10.15698/cst2020.04.216
- de Wit E, van Doremalen N, Falzarano D, and Munster VJ (2016). SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol 14(8): 523–534. doi: 10.1038/nrmicro.2016.81
- Ye Z-W, Yuan S, Yuen K-S, Fung S-Y, Chan C-P, and Jin D-Y (2020). Zoonotic origins of human coronaviruses. Int J Biol Sci 16(10): 1686–1697. doi: 10.7150/ijbs.45472
- Yin Z, Pascual C, and Klionsky DJ (2016). Autophagy: machinery and regulation. Microb Cell 3(12): 588–596. doi: 10.15698/mic2016.12.546
- Eisenberg T, Knauer H, Schauer A, Büttner S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes C, Antonacci L, Fussi H, Deszcz L, Hartl R, Schraml E, Criollo A, Megalou E, Weiskopf D, Laun P, Heeren G, Breitenbach M, Grubeck-Loebenstein B, Herker E, Fahrenkrog B, Fröhlich K-U, Sinner F, Tavernarakis N, Minois N, Kroemer G, and Madeo F (2009). Induction of autophagy by spermidine promotes longevity. Nat Cell Biol 11(11): 1305–1314. doi: 10.1038/ncb1975
- Carmona-Gutierrez D et al. (2019). The flavonoid 4,4′-dimethoxychalcone promotes autophagy-dependent longevity across species. Nat Commun 10(1): 651. doi: 10.1038/s41467-019-08555-w
- Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, and Sinclair DA (2006). Resveratrol improves health and survival of mice on a high-calorie diet. Nature 444(7117): 337–342. doi: 10.1038/nature05354
- Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, and Miller RA (2009). Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 460(7253): 392–395. doi: 10.1038/nature08221
- Chen G, Xie W, Nah J, Sauvat A, Liu P, Pietrocola F, Sica V, Carmona-Gutierrez D, Zimmermann A, Pendl T, Tadic J, Bergmann M, Hofer SJ, Domuz L, Lachkar S, Markaki M, Tavernarakis N, Sadoshima J, Madeo F, Kepp O, and Kroemer G (2019). 3,4-Dimethoxychalcone induces autophagy through activation of the transcription factors TFE3 and TFEB. EMBO Mol Med 11(11): e10469. doi: 10.15252/emmm.201910469
- de Cabo R, Carmona-Gutierrez D, Bernier M, Hall MN, and Madeo F (2014). The search for antiaging interventions: from elixirs to fasting regimens. Cell 157(7): 1515–1526. doi: 10.1016/j.cell.2014.05.031
- Levine B, and Kroemer G (2008). Autophagy in the Pathogenesis of Disease. Cell 132(1): 27–42. doi: 10.1016/j.cell.2007.12.018
- Kirkin V, and Rogov VV (2019). A Diversity of Selective Autophagy Receptors Determines the Specificity of the Autophagy Pathway. Mol Cell 76(2): 268–285. doi: 10.1016/j.molcel.2019.09.005
- Stolz A, Ernst A, and Dikic I (2014). Cargo recognition and trafficking in selective autophagy. Nat Cell Biol 16(6): 495–501. doi: 10.1038/ncb2979
- Gatica D, Lahiri V, and Klionsky DJ (2018). Cargo recognition and degradation by selective autophagy. Nat Cell Biol 20(3): 233–242. doi: 10.1038/s41556-018-0037-z
- Sharma V, Verma S, Seranova E, Sarkar S, and Kumar D (2018). Selective Autophagy and Xenophagy in Infection and Disease. Front Cell Dev Biol 6: 147. doi: 10.3389/fcell.2018.00147
- Sagnier S, Daussy CF, Borel S, Robert-Hebmann V, Faure M, Blanchet FP, Beaumelle B, Biard-Piechaczyk M, and Espert L (2015). Autophagy restricts HIV-1 infection by selectively degrading Tat in CD4+ T lymphocytes. J Virol 89(1): 615–625. doi: 10.1128/JVI.02174-14
- Nardacci R, Amendola A, Ciccosanti F, Corazzari M, Esposito V, Vlassi C, Taibi C, Fimia GM, Del Nonno F, Ippolito G, D’Offizi G, and Piacentini M (2014). Autophagy plays an important role in the containment of HIV-1 in nonprogressor-infected patients. Autophagy 10(7): 1167–1178. doi: 10.4161/auto.28678
- Sparrer KMJ, Gableske S, Zurenski MA, Parker ZM, Full F, Baumgart GJ, Kato J, Pacheco-Rodriguez G, Liang C, Pornillos O, Moss J, Vaughan M, and Gack MU (2017). TRIM23 mediates virus-induced autophagy via activation of TBK1. Nat Microbiol 2(11): 1543–1557. doi: 10.1038/s41564-017-0017-2
- Yordy B, Iijima N, Huttner A, Leib D, and Iwasaki A (2012). A neuron-specific role for autophagy in antiviral defense against herpes simplex virus. Cell Host Microbe 12(3): 334–345. doi: 10.1016/j.chom.2012.07.013
- Choi Y, Bowman JW, and Jung JU (2018). Autophagy during viral infection — a double-edged sword. Nat Rev Microbiol 16(6): 341–354. doi: 10.1038/s41579-018-0003-6
- Cong Y, Verlhac P, and Reggiori F (2017). The Interaction between Nidovirales and Autophagy Components. Viruses 9(7): 182. doi: 10.3390/v9070182
- Shi C-S, Nabar NR, Huang N-N, and Kehrl JH (2019). SARS-Coronavirus Open Reading Frame-8b triggers intracellular stress pathways and activates NLRP3 inflammasomes. Cell Death Discov 5: 101. doi: 10.1038/s41420-019-0181-7
- Yue Y, Nabar NR, Shi C-S, Kamenyeva O, Xiao X, Hwang I-Y, Wang M, and Kehrl JH (2018). SARS-Coronavirus Open Reading Frame-3a drives multimodal necrotic cell death. Cell Death Dis 9(9): 904. doi: 10.1038/s41419-018-0917-y
- Gassen NC, Niemeyer D, Muth D, Corman VM, Martinelli S, Gassen A, Hafner K, Papies J, Mösbauer K, Zellner A, Zannas AS, Herrmann A, Holsboer F, Brack-Werner R, Boshart M, Müller-Myhsok B, Drosten C, Müller MA, and Rein T (2019). SKP2 attenuates autophagy through Beclin1-ubiquitination and its inhibition reduces MERS-Coronavirus infection. Nat Commun 10. doi: 10.1038/s41467-019-13659-4
- Jackson WT (2015). Viruses and the Autophagy Pathway. Virology 479–480: 450–456. doi: 10.1016/j.virol.2015.03.042
- Papandreou M-E, and Tavernarakis N (2017). Autophagy and the endo/exosomal pathways in health and disease. Biotechnol J 12(1): 1600175. doi: 10.1002/biot.201600175
- Gosert R, Kanjanahaluethai A, Egger D, Bienz K, and Baker SC (2002). RNA replication of mouse hepatitis virus takes place at double-membrane vesicles. J Virol 76(8): 3697–3708. doi: 10.1128/jvi.76.8.3697-3708.2002
- Lundin A, Dijkman R, Bergström T, Kann N, Adamiak B, Hannoun C, Kindler E, Jónsdóttir HR, Muth D, Kint J, Forlenza M, Müller MA, Drosten C, Thiel V, and Trybala E (2014). Targeting membrane-bound viral RNA synthesis reveals potent inhibition of diverse coronaviruses including the middle East respiratory syndrome virus. PLoS Pathog 10(5): e1004166. doi: 10.1371/journal.ppat.1004166
- Knoops K, Kikkert M, Worm SHE van den, Zevenhoven-Dobbe JC, van der Meer Y, Koster AJ, Mommaas AM, and Snijder EJ (2008). SARS-coronavirus replication is supported by a reticulovesicular network of modified endoplasmic reticulum. PLoS Biol 6(9): e226. doi: 10.1371/journal.pbio.0060226
- Prentice E, Jerome WG, Yoshimori T, Mizushima N, and Denison MR (2004). Coronavirus replication complex formation utilizes components of cellular autophagy. J Biol Chem 279(11): 10136–10141. doi: 10.1074/jbc.M306124200
- Prentice E, McAuliffe J, Lu X, Subbarao K, and Denison MR (2004). Identification and Characterization of Severe Acute Respiratory Syndrome Coronavirus Replicase Proteins. J Virol 78(18): 9977–9986. doi: 10.1128/JVI.78.18.9977-9986.2004
- Zhu L, Mou C, Yang X, Lin J, and Yang Q (2016). Mitophagy in TGEV infection counteracts oxidative stress and apoptosis. Oncotarget 7(19): 27122–27141. doi: 10.18632/oncotarget.8345
- Guo X, Zhang M, Zhang X, Tan X, Guo H, Zeng W, Yan G, Memon AM, Li Z, Zhu Y, Zhang B, Ku X, Wu M, Fan S, and He Q (2017). Porcine Epidemic Diarrhea Virus Induces Autophagy to Benefit Its Replication. Viruses 9(3): E53. doi: 10.3390/v9030053
- Reggiori F, Monastyrska I, Verheije MH, Calì T, Ulasli M, Bianchi S, Bernasconi R, de Haan CAM, and Molinari M (2010). Coronaviruses Hijack the LC3-I-positive EDEMosomes, ER-derived vesicles exporting short-lived ERAD regulators, for replication. Cell Host Microbe 7(6): 500–508. doi: 10.1016/j.chom.2010.05.013
- Zhao Z, Thackray LB, Miller BC, Lynn TM, Becker MM, Ward E, Mizushima NN, Denison MR, and Virgin HW (2007). Coronavirus replication does not require the autophagy gene ATG5. Autophagy 3(6): 581–585. doi: 10.4161/auto.4782
- Schneider M, Ackermann K, Stuart M, Wex C, Protzer U, Schätzl HM, and Gilch S (2012). Severe acute respiratory syndrome coronavirus replication is severely impaired by MG132 due to proteasome-independent inhibition of M-calpain. J Virol 86(18): 10112–10122. doi: 10.1128/JVI.01001-12
- Snijder EJ, van der Meer Y, Zevenhoven-Dobbe J, Onderwater JJM, van der Meulen J, Koerten HK, and Mommaas AM (2006). Ultrastructure and Origin of Membrane Vesicles Associated with the Severe Acute Respiratory Syndrome Coronavirus Replication Complex. J Virol 80(12): 5927–5940. doi: 10.1128/JVI.02501-05
- Guo L, Yu H, Gu W, Luo X, Li R, Zhang J, Xu Y, Yang L, Shen N, Feng L, and Wang Y (2016). Autophagy Negatively Regulates Transmissible Gastroenteritis Virus Replication. Sci Rep 6: 23864. doi: 10.1038/srep23864
- Ko S, Gu MJ, Kim CG, Kye YC, Lim Y, Lee JE, Park B-C, Chu H, Han SH, and Yun C-H (2017). Rapamycin-induced autophagy restricts porcine epidemic diarrhea virus infectivity in porcine intestinal epithelial cells. Antiviral Res 146: 86–95. doi: 10.1016/j.antiviral.2017.08.010
- Dong X, and Levine B (2013). Autophagy and viruses: adversaries or allies? J Innate Immun 5(5): 480–493. doi: 10.1159/000346388
- Heckmann BL, and Green DR (2019). LC3-associated phagocytosis at a glance. J Cell Sci 132(5). doi: 10.1242/jcs.222984
- Grose C, and Klionsky DJ (2016). Alternative autophagy, brefeldin A and viral trafficking pathways. Autophagy 12(9): 1429–1430. doi: 10.1080/15548627.2016.1203489
- Nishida Y, Arakawa S, Fujitani K, Yamaguchi H, Mizuta T, Kanaseki T, Komatsu M, Otsu K, Tsujimoto Y, and Shimizu S (2009). Discovery of Atg5/Atg7-independent alternative macroautophagy. Nature 461(7264): 654–658. doi: 10.1038/nature08455
- Niso-Santano M, Malik SA, Pietrocola F, Bravo-San Pedro JM, Mariño G, Cianfanelli V, Ben-Younès A, Troncoso R, Markaki M, Sica V, Izzo V, Chaba K, Bauvy C, Dupont N, Kepp O, Rockenfeller P, Wolinski H, Madeo F, Lavandero S, Codogno P, Harper F, Pierron G, Tavernarakis N, Cecconi F, Maiuri MC, Galluzzi L, and Kroemer G (2015). Unsaturated fatty acids induce non-canonical autophagy. EMBO J.34(8): 1025–1041. doi: 10.15252/embj.201489363
- Kindrachuk J, Ork B, Hart BJ, Mazur S, Holbrook MR, Frieman MB, Traynor D, Johnson RF, Dyall J, Kuhn JH, Olinger GG, Hensley LE, and Jahrling PB (2015). Antiviral potential of ERK/MAPK and PI3K/AKT/mTOR signaling modulation for Middle East respiratory syndrome coronavirus infection as identified by temporal kinome analysis. Antimicrob Agents Chemother 59(2): 1088–1099. doi: 10.1128/AAC.03659-14
- Cottam EM, Maier HJ, Manifava M, Vaux LC, Chandra-Schoenfelder P, Gerner W, Britton P, Ktistakis NT, and Wileman T (2011). Coronavirus nsp6 proteins generate autophagosomes from the endoplasmic reticulum via an omegasome intermediate. Autophagy 7(11): 1335–1347. doi: 10.4161/auto.7.11.16642
- Cottam EM, Whelband MC, and Wileman T (2014). Coronavirus NSP6 restricts autophagosome expansion. Autophagy 10(8): 1426–1441. doi: 10.4161/auto.29309
- Chan JF-W, Kok K-H, Zhu Z, Chu H, To KK-W, Yuan S, and Yuen K-Y (2020). Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg Microbes Infect 9(1): 221–236. doi: 10.1080/22221751.2020.1719902
- Chen X, Wang K, Xing Y, Tu J, Yang X, Zhao Q, Li K, and Chen Z (2014). Coronavirus membrane-associated papain-like proteases induce autophagy through interacting with Beclin1 to negatively regulate antiviral innate immunity. Protein Cell 5(12): 912–927. doi: 10.1007/s13238-014-0104-6
- Gusho E, Baskar D, and Banerjee S (2016). New advances in our understanding of the “unique” RNase L in host pathogen interaction and immune signaling. Cytokine. doi: 10.1016/j.cyto.2016.08.009
- Yang Y, Zhang L, Geng H, Deng Y, Huang B, Guo Y, Zhao Z, and Tan W (2013). The structural and accessory proteins M, ORF 4a, ORF 4b, and ORF 5 of Middle East respiratory syndrome coronavirus (MERS-CoV) are potent interferon antagonists. Protein Cell 4(12): 951–961. doi: 10.1007/s13238-013-3096-8
- Ambjørn M, Ejlerskov P, Liu Y, Lees M, Jäättelä M, and Issazadeh-Navikas S (2013). IFNB1/interferon-β-induced autophagy in MCF-7 breast cancer cells counteracts its proapoptotic function. Autophagy 9(3): 287–302. doi: 10.4161/auto.22831
- Muth D, Corman VM, Roth H, Binger T, Dijkman R, Gottula LT, Gloza-Rausch F, Balboni A, Battilani M, Rihtarič D, Toplak I, Ameneiros RS, Pfeifer A, Thiel V, Drexler JF, Müller MA, and Drosten C (2018). Attenuation of replication by a 29 nucleotide deletion in SARS-coronavirus acquired during the early stages of human-to-human transmission. Sci Rep 8(1): 15177. doi: 10.1038/s41598-018-33487-8
- Liu DX, Fung TS, Chong KK-L, Shukla A, and Hilgenfeld R (2014). Accessory proteins of SARS-CoV and other coronaviruses. Antiviral Res 109: 97–109. doi: 10.1016/j.antiviral.2014.06.013
- Vincent MJ, Bergeron E, Benjannet S, Erickson BR, Rollin PE, Ksiazek TG, Seidah NG, and Nichol ST (2005). Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol J 2: 69. doi: 10.1186/1743-422X-2-69
- Gautret P, Lagier J-C, Parola P, Hoang VT, Meddeb L, Mailhe M, Doudier B, Courjon J, Giordanengo V, Vieira VE, Dupont HT, Honoré S, Colson P, Chabrière E, La Scola B, Rolain J-M, Brouqui P, and Raoult D (2020). Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial. Int J Antimicrob Agents 105949. doi: 10.1016/j.ijantimicag.2020.105949
- Wang M, Cao R, Zhang L, Yang X, Liu J, Xu M, Shi Z, Hu Z, Zhong W, and Xiao G (2020). Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res 30(3): 269–271. doi: 10.1038/s41422-020-0282-0
- Jaffe S (2020). Regulators split on antimalarials for COVID-19. The Lancet 395(10231): 1179. doi: 10.1016/S0140-6736(20)30817-5
- Ferner RE, and Aronson JK (2020). Chloroquine and hydroxychloroquine in covid-19. BMJ 369: m1432. doi: 10.1136/bmj.m1432
- Gbinigie K, and Frie K (2020). Should chloroquine and hydroxychloroquine be used to treat COVID-19? A rapid review. BJGP Open. doi: 10.3399/bjgpopen20X101069
- Mauthe M, Orhon I, Rocchi C, Zhou X, Luhr M, Hijlkema K-J, Coppes RP, Engedal N, Mari M, and Reggiori F (2018). Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 14(8): 1435–1455. doi: 10.1080/15548627.2018.1474314
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu N-H, Nitsche A, Müller MA, Drosten C, and Pöhlmann S (2020). SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181(2): 271-280.e8. doi: 10.1016/j.cell.2020.02.052
- Al-Bari MdAA (2017). Targeting endosomal acidification by chloroquine analogs as a promising strategy for the treatment of emerging viral diseases. Pharmacol Res Perspect 5(1): e00293. doi: 10.1002/prp2.293
- Walls AC, Park Y-J, Tortorici MA, Wall A, McGuire AT, and Veesler D (2020). Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 181(2):281-292.e6. doi: 10.1016/j.cell.2020.02.058
- Zhao X, Guo F, Comunale MA, Mehta A, Sehgal M, Jain P, Cuconati A, Lin H, Block TM, Chang J, and Guo J-T (2015). Inhibition of endoplasmic reticulum-resident glucosidases impairs severe acute respiratory syndrome coronavirus and human coronavirus NL63 spike protein-mediated entry by altering the glycan processing of angiotensin I-converting enzyme 2. Antimicrob Agents Chemother 59(1): 206–216. doi: 10.1128/AAC.03999-14
- Wu K, Zhang Q, Wu X, Lu W, Tang H, Liang Z, Gu Y, Song S, Ayon RJ, Wang Z, McDermott KM, Balistrieri A, Wang C, Black SM, Garcia JGN, Makino A, Yuan JX-J, and Wang J (2017). Chloroquine is a potent pulmonary vasodilator that attenuates hypoxia-induced pulmonary hypertension. Br J Pharmacol 174(22): 4155–4172. doi: 10.1111/bph.13990
- Martina JA, Diab HI, Lishu L, Jeong-A L, Patange S, Raben N, and Puertollano R (2014). The Nutrient-Responsive Transcription Factor TFE3, Promotes Autophagy, Lysosomal Biogenesis, and Clearance of Cellular Debris. Sci Signal 7(309): ra9. doi: 10.1126/scisignal.2004754
- Roczniak-Ferguson A, Petit CS, Froehlich F, Qian S, Ky J, Angarola B, Walther TC, and Ferguson SM (2012). The transcription factor TFEB links mTORC1 signaling to transcriptional control of lysosome homeostasis. Sci Signal 5(228): ra42. doi: 10.1126/scisignal.2002790
- Gordon DE et al. (2020). A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug-Repurposing. bioRxiv [Preprint]. Version 3, posted March 27, 2020. doi: 10.1101/2020.03.22.002386
- Le Sage V, Cinti A, Amorim R, and Mouland AJ (2016). Adapting the Stress Response: Viral Subversion of the mTOR Signaling Pathway. Viruses 8(6): 152. doi: 10.3390/v8060152
- Ranadheera C, Coombs KM, and Kobasa D (2018). Comprehending a Killer: The Akt/mTOR Signaling Pathways Are Temporally High-Jacked by the Highly Pathogenic 1918 Influenza Virus. EBioMedicine 32: 142–163. doi: 10.1016/j.ebiom.2018.05.027
- Gassen NC, Papies J, Bajaj T, Dethloff F, Emanuel J, Weckmann K, Heinz DE, Heinemann N, Lennarz M, Richter A, Niemeyer D, Corman VM, Giavalisco P, Drosten C, and Muller MA (2020). Analysis of SARS-CoV-2-controlled autophagy reveals spermidine, MK-2206, and niclosamide as putative antiviral therapeutics. bioRxiv [Preprint]. Version 1, posted April 15, 2020. doi: 10.1101/2020.04.15.997254
- Pegg AE (2016). Functions of Polyamines in Mammals. J Biol Chem 291(29): 14904–14912. doi: 10.1074/jbc.R116.731661
- Madeo F, Eisenberg T, Pietrocola F, and Kroemer G (2018). Spermidine in health and disease. Science 359(6374): eaan2788. doi: 10.1126/science.aan2788
- Eisenberg T et al. (2016). Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat Med 22(12): 1428–1438. doi: 10.1038/nm.4222
- Morselli E, Mariño G, Bennetzen MV, Eisenberg T, Megalou E, Schroeder S, Cabrera S, Bénit P, Rustin P, Criollo A, Kepp O, Galluzzi L, Shen S, Malik SA, Maiuri MC, Horio Y, López-Otín C, Andersen JS, Tavernarakis N, Madeo F, and Kroemer G (2011). Spermidine and resveratrol induce autophagy by distinct pathways converging on the acetylproteome. J Cell Biol 192(4): 615–629. doi: 10.1083/jcb.201008167
- Gupta VK, Scheunemann L, Eisenberg T, Mertel S, Bhukel A, Koemans TS, Kramer JM, Liu KSY, Schroeder S, Stunnenberg HG, Sinner F, Magnes C, Pieber TR, Dipt S, Fiala A, Schenck A, Schwaerzel M, Madeo F, and Sigrist SJ (2013). Restoring polyamines protects from age-induced memory impairment in an autophagy-dependent manner. Nat Neurosci 16(10): 1453–1460. doi: 10.1038/nn.3512
- Sonawane ND, Szoka FC, and Verkman AS (2003). Chloride accumulation and swelling in endosomes enhances DNA transfer by polyamine-DNA polyplexes. J Biol Chem 278(45): 44826–44831. doi: 10.1074/jbc.M308643200
- Mounce BC, Olsen ME, Vignuzzi M, and Connor JH (2017). Polyamines and Their Role in Virus Infection. Microbiol Mol Biol Rev 81(4): e00029-17. doi: 10.1128/MMBR.00029-17
- Mounce BC, Poirier EZ, Passoni G, Simon-Loriere E, Cesaro T, Prot M, Stapleford KA, Moratorio G, Sakuntabhai A, Levraud J-P, and Vignuzzi M (2016). Interferon-Induced Spermidine-Spermine Acetyltransferase and Polyamine Depletion Restrict Zika and Chikungunya Viruses. Cell Host Microbe 20(2): 167–177. doi: 10.1016/j.chom.2016.06.011
- Zhu Y, Deng J, Nan M-L, Zhang J, Okekunle A, Li J-Y, Yu X-Q, and Wang P-H (2019). The Interplay Between Pattern Recognition Receptors and Autophagy in Inflammation. Adv Exp Med Biol 1209: 79–108. doi: 10.1007/978-981-15-0606-2_6
- Akira S, Uematsu S, and Takeuchi O (2006). Pathogen recognition and innate immunity. Cell 124(4): 783–801. doi: 10.1016/j.cell.2006.02.015
- Dengjel J, Schoor O, Fischer R, Reich M, Kraus M, Müller M, Kreymborg K, Altenberend F, Brandenburg J, Kalbacher H, Brock R, Driessen C, Rammensee H-G, Stevanovic S, and Cresswell P (2005). Autophagy Promotes MHC Class II Presentation of Peptides from Intracellular Source Proteins. Proc Natl Acad Sci U S A 102(22): 7922–7927. doi: 10.1073/pnas.0501190102
- Münz C (2016). Autophagy Beyond Intracellular MHC Class II Antigen Presentation. Trends Immunol 37(11): 755–763. doi: 10.1016/j.it.2016.08.017
- English L, Chemali M, Duron J, Rondeau C, Laplante A, Gingras D, Alexander D, Leib D, Norbury C, Lippé R, and Desjardins M (2009). Autophagy enhances the presentation of endogenous viral antigens on MHC class I molecules during HSV-1 infection. Nat Immunol 10(5): 480–487. doi: 10.1038/ni.1720
- Jiang G-M, Tan Y, Wang H, Peng L, Chen H-T, Meng X-J, Li L-L, Liu Y, Li W-F, and Shan H (2019). The relationship between autophagy and the immune system and its applications for tumor immunotherapy. Mol Cancer 18(1): 17. doi: 10.1186/s12943-019-0944-z
- Pietrocola F et al. (2016). Caloric Restriction Mimetics Enhance Anticancer Immunosurveillance. Cancer Cell 30(1): 147–160. doi: 10.1016/j.ccell.2016.05.016
- Puleston DJ, Zhang H, Powell TJ, Lipina E, Sims S, Panse I, Watson AS, Cerundolo V, Townsend AR, Klenerman P, and Simon AK (2014). Autophagy is a critical regulator of memory CD8+ T cell formation. eLife 3. doi: 10.7554/eLife.03706
- Zhang H, Alsaleh G, Feltham J, Sun Y, Napolitano G, Riffelmacher T, Charles P, Frau L, Hublitz P, Yu Z, Mohammed S, Ballabio A, Balabanov S, Mellor J, and Simon AK (2019). Polyamines Control eIF5A Hypusination, TFEB Translation, and Autophagy to Reverse B Cell Senescence. Mol Cell 76(1): 110-125.e9. doi: 10.1016/j.molcel.2019.08.005
- Harris J, Lang T, Thomas JPW, Sukkar MB, Nabar NR, and Kehrl JH (2017). Autophagy and inflammasomes. Mol Immunol 86: 10–15. doi: 10.1016/j.molimm.2017.02.013
- Ponpuak M, Mandell MA, Kimura T, Chauhan S, Cleyrat C, and Deretic V (2015). Secretory autophagy. Curr Opin Cell Biol 35: 106–116. doi: 10.1016/j.ceb.2015.04.016
- Delorme-Axford E, Donker RB, Mouillet J-F, Chu T, Bayer A, Ouyang Y, Wang T, Stolz DB, Sarkar SN, Morelli AE, Sadovsky Y, and Coyne CB (2013). Human placental trophoblasts confer viral resistance to recipient cells. Proc Natl Acad Sci U S A 110(29): 12048–12053. doi: 10.1073/pnas.1304718110
- Ahmad L, Mashbat B, Leung C, Brookes C, Hamad S, Krokowski S, Shenoy AR, Lorenzo L, Levin M, O’Hare P, Zhang S-Y, Casanova J-L, Mostowy S, and Sancho-Shimizu V (2019). Human TANK-binding kinase 1 is required for early autophagy induction upon herpes simplex virus 1 infection. J Allergy Clin Immunol 143(2): 765-769.e7. doi: 10.1016/j.jaci.2018.09.013
- Kepp O, Chen G, Carmona-Gutierrez D, Madeo F, and Kroemer G (2020). A discovery platform for the identification of caloric restriction mimetics with broad health-improving effects. Autophagy 16(1): 188–189. doi: 10.1080/15548627.2019.1688984
- Madeo F, Carmona-Gutierrez D, Hofer SJ, and Kroemer G (2019). Caloric Restriction Mimetics against Age-Associated Disease: Targets, Mechanisms, and Therapeutic Potential. Cell Metab 29(3): 592–610. doi: 10.1016/j.cmet.2019.01.018
ACKNOWLEDGMENTS
The authors are grateful to the Aus-trian Science Fund FWF (SFBLIPOTOX F3007&F3012, W1226, P29203, P29262, P27893) and the Austrian Federal Ministry of Education, Science and Research and the University of Graz for grants “Unkonventionelle For-schung” and “flysleep” (BMWFW-80.109/0001-WF/V/3b/2015). The authors also acknowledge the funding of DK Metabolic and Cardiovascular Disease (FWF) and the Doctoral College “Metabolic and Cardio-vascular Disease” (FWFW1226) as well as support from NAWI Graz and the BioTechMed-Graz flagship project “EPIAge”. GK is supported by the Ligue contre le Cancer (équipe labellisée); Agence National de la Recherche (ANR) – Projets blancs; ANR under the frame of E-Rare-2, the ERA-Net for Research on Rare Diseases; AMMICa US23/CNRS UMS3655; Association pour la recherche sur le cancer (ARC); Association "Le Cancer du Sein, Parlons-en!"; Cancéropôle Ile-de-France; Chancelerie des uni-versités de Paris (Legs Poix), Fondation pour la Recher-che Médicale (FRM); a dona-tion by Elior; European Re-search Area Network on Car-diovascular Diseases (ERA-CVD, MINOTAUR); Gustave Roussy Odyssea, the Europe-an Union Horizon 2020 Pro-ject Oncobiome; Fondation Carrefour; High-end Foreign Expert Program in China (GDW20171100085), Institut National du Cancer (INCa); Inserm (HTE); Institut Univer-sitaire de France; LeDucq Foundation; the LabEx Immu-no-Oncology (ANR-18-IDEX-0001); the RHU Torino Lu-mière; the Seerave Founda-tion; the SIRIC Stratified On-cology Cell DNA Repair and Tumor Immune Elimination (SOCRATE); and the SIRIC Cancer Research and Personalized Medicine (CARPEM).
COPYRIGHT
© 2020

Digesting the crisis: autophagy and coronaviruses by Carmona-Gutierrez et al. is licensed under a Creative Commons Attribution 4.0 International License.








